

自闭症谱系障碍(ASD)等神经发育障碍中存在大量15q11.2-q13.1染色体倍增(Dup15q综合征,Duplications of 15q syndrome)个体,该染色体包括父系印记基因UBE3A和三个非印记γ-氨基丁酸(GABAA)受体基因。研究发现Dup15q综合征存在典型β-EEG活动,但不清楚是由哪个特定基因表达。
为了验证UBE3A过度表达并不一定产生β-EEG活动,本研究将Dup15q综合征儿童(研究组)与以下几组进行比较:
(1)使用GABAA调节剂咪达唑仑(midazolam)的健康成人
(2)正常(TD)儿童
(3)两个父系倍增15q(即UBE3A沉默等位基因)儿童。
结果发现,与TD相比,研究组β峰值功率显著增加。咪达唑仑诱发的健康成人中线电极β-EEG活动存在与Dup15q综合征相似的峰值频率。两个父系倍增儿童β功率与研究组相似。
研究结果说明,不能单用UBE3A功能障碍来解释 Dup15q综合征β-EEG活动,GABAergic对β-EEG活动有关键作用。且β-EEG活动可作为Dup15q综合征临床试验中GABAergic病理学的标记物,指导Dup15q综合征和其他形式ASD的靶向治疗。本文发表在Molecular Autism杂志。
关键词:Dup15q syndrome, GABA, UBE3A, Biomarkers, Autism, EEG, Neurodevelopmental disorders, GABRA5, GABRB3, GABRG3
研究背景
15q11.2-q13.1(Dup15q综合征)染色体倍增和三倍复制(triplications)大量出现在智力障碍(ID,intellectual disability)、自闭症谱系障碍(ASD)、发育迟缓和癫痫中,是ASD最常见的基因拷贝数变异(CNV,copy number variant)。该区域几个基因影响大脑突触功能和抑制性神经传递的早期发育。通过分别检测母系和父系染色体倍增情况,阐明等位基因在神经元中的特异性表达(即母系或父系印记),从而研究这些基因对Dup15q的病理性作用。
Dup15q综合征临床症状受亲代印记影响,母系倍增儿童表现出更严重的临床症状和更大的ASD和ID的可能性。这种差异可能是因为编码泛素蛋白连接酶并调节突触发育的父系印记UBE3A基因与神经发育障碍有关。存在两种15q倍增类型:间质(interstitial)倍增和等双中心(isodicentric)倍增。间质倍增表现为15号染色体内15q11.2-q13.1额外拷贝,这会导致部分三体或少见的部分四体。等双中心倍增是15q11.2-q13.1端到端连接一个额外染色体的额外拷贝,导致部分四体,并产生更严重的临床症状。50%以上母系间质和80%以上等双中心倍增的儿童符合ASD诊断标准。 Dup15q综合征和其他形式ASD的靶向治疗中需要可以反映治疗效果的分子或神经环路水平的生物标记物。这种基于生物机制的标记物可以作为检测持续时间短且无法观察长期行为变化的临床试验有效性的替代参考指标,也可以作为药物靶点参与的量化指标,反过来又可以为试验进行提供决策信息。Dup15q综合征具有可能反映分子病理的典型β-EEG活动。未服用苯二氮卓类或其他已知可诱发β活动的药物的Dup15q综合征儿童的β-EEG活动可量化为自发β(12-30Hz)振荡。因此,β-EEG活动是Dup15q综合征治疗发展中病理生理或药物靶点的可量化生物标记物。
正确应用Dup15q综合征标记物的关键取决于了解该生物标记物反映哪些基因和病理生理学的哪些方面。某些15q染色体基因,包括父系印记基因UBE3A和一组非印记γ-氨基丁酸(GABAA)受体β3、α5和γ3亚基,都与疾病病源有关。本研究为了推断UBE3A或GABRB3/GABRA5/GABRG3基因簇对Dup15q综合征β活动的必要性或充分性,进行了三项研究。
1)为了证实Dup15q综合征更大样本中仍然存在β-EEG活动,将正常儿童的β活动与具有间质和等双中心倍增Dup15q综合征儿童的β活动进行了比较。
2)为了验证GABAergic功能障碍足以产生β-EEG活动的假设,将Dup15q综合征β-EEG活动与通过GABAA调节剂咪唑安定药物诱发的健康成人β活动进行了比较。
3)为了验证UBE3A失调对β-EEG活动的必要性,将两例父系Dup15q综合征与上述Dup15q综合征儿童进行比较。
方法
被试招募和与脑电数据采集
本研究包含以下三类被试:
(1)27名Dup15q综合征儿童(研究组)
(2)14名正常儿童(TD组)
(3)两名父系15q11-q13倍增儿童
(4)12名使用5毫克咪唑安定的健康成人志愿者。
(5)两名父系Dup15q综合征儿童的数据来自先前研究中的临床脑电。
数据采集的详细情况如下:
Dup15q综合征儿童组
与患者权益团体合作,在全国性家庭会议上招募儿童。招募所有年龄和发育能力的儿童来获取最具临床代表性的样本。研究开始之前被试父母签署知情同意书。谨慎排除具有混杂因素的被试数据,如癫痫和使用对GABAergic传递起作用的抗癫痫药物的被试。此外,研究组排除父系倍增儿童。多数遗传报告中不包含亲本数据,因此只确认了两例母系亲本案例。不过研究组中绝大多数可能是母系倍增儿童,因为母系倍增的情况大约是父系倍增的两倍,在ASD和发育迟缓中大约是2.5倍。研究组共27名Dup15q综合征患者(13名间质,14名等中心),年龄和发育商(DQ)详情见表1。
使用129导Ag/AgCl以顶点为参考,500Hz采样率采集高密度(HD,High density)脑电数据。
TD组
为了对照确认Dup15q研究组独特的β高功率,还检测了14名TD儿童清醒状态自发脑电数据。两组被试年龄无显著差异(p=0.29)。招募、知情同意和脑电图方案与上述Dup15q研究组相同,年龄和DQ详情见表1。
表1 Dup15q综合征儿童组参与者详细信息注:
DQ(developmental quotient)为发育商数。
父系Dup15q综合征病例组
父系Dup15q综合征临床表现温和,其诊断率比母系Dup15q综合征要低,因此本研究只采集两个来自不同研究机构的父系倍增儿童的脑电数据(见表2)。这两个父系倍增被试都参加过一项间质Dup15q综合征研究。因此,本研究直接使用已有编码。第一位被试(801-005)是一名13岁(161个月)患有父系间质Dup15q综合征的男孩,通过高密度EGI收集其清醒状态自发脑电数据。第二个被试(801-015)是一名8岁(96个月)患有父系间质Dup15q综合征的女孩,收集清醒状态临床自发脑电图数据(采样率=512Hz)。使用平均参考对数据进行重设参考,对除双耳之外的19个电极数据(标准10-20系统)进行分析。
表2 父系Dup15q综合征病例组参与者详细信息及其脑电信息

咪达唑仑药物诱发脑电组
为了评估Dup15q综合征的β活动与GABAA异构激活调节剂(PAM)诱发的β活动的相似性,收集12名接受苯二氮类化合物咪唑安定的健康成人19导脑电数据(采样率=256Hz)。咪达唑仑是一种非选择性的GABAA-PAM(比如,苯二氮卓),它与GABAA受体结合打开离子通道,增加受体效能。
脑电数据预处理
通过MATLAB进行数据处理。数据经过1-45Hz带通滤波(FIR滤波器),使用人工伪迹判断和独立成分分析(ICA)减少伪迹。从高密度脑电数据中去除对噪声和肌肉伪迹特别敏感的46个“外周电极”(skirt channels),剩下83个电极用于后续处理和分析。通过人工伪迹判断去除系统噪音和大肌肉运动伪迹,噪声电极被标记为插值,并且排除噪声电极数超过总电极数平方根的被试数据。每个被试至少有60秒的干净数据进行后续分析。采用FastICA算法进行ICA。从脑电图数据中去除生理伪迹(如眨眼、眼跳、颈部运动)相对应的成分。在去除伪迹后,对坏电极进行插值替换。小波变换前以平均参照进行重设参考。由于直接比较HD-EEG和19-EEG的数据,在小波变换之前将HD-EEG对应为10-20分布的19电极。
频域变换与分析
用Morlet小波进行频域变换。共使用54个Morlet小波核,对数间隔频率为2-45Hz(每倍频有12个小波),频谱平滑度为1/3倍频。接下来,删除已排除数据的时间窗对应的时频成分。去除有效(即非排除的)数据中2Hz小波变换时间窗口少于20的数据。通过计算干净数据中连续3/4重叠时间窗的平均功率值得到频谱功率值,共得到83个(HD-EEG)或19个(10-20系统)电极和54个频率中每个电极的谱功率。 使用半倍频平滑频率输出,并用log2(Hz)(比如,倍频)对每个单位的值进行标准化从而得到功率谱密度(PSD)。然后,使用log2(Hz)计算PSDs,并在对数空间中绘制PSDs,以解释电生理信号的对数性质。由于相对功率测量容易受到标准化伪迹的影响,后续计算中使用绝对功率。
为了比较父系Dup15q综合征病例与研究组的PSDs,使用线性回归来解释年龄差异。由于对数转换后可以解释更年轻时的更大发展变化,使用log2(年龄)为研究组中的所有被试建立了PSD模型。然后,使用父系Dup15q综合征被试的log2(年龄)重建每个研究组被试的PSD,并为每个被试添加反向模型残差。
峰值频率提取
为了进一步研究β活动,本研究计算Dup15q综合征(研究组和父系倍增的被试)的峰值β。在额叶电极的平均功率上计算峰值β,因为Dup15q综合征该区域的β功率最高(图1)。使用插值方法将HD EEG在空间上对应到国际10-20导联中的19个电极,并且平均电极Fp1、Fp2、F3、F4、Fz、F7和F8的功率。通过提取出现最大功率值的β频率,在每个被试中自动识别额叶峰值所在的β频率(FBPF,Frontal beta peak frequency)。
结果
Dup15q研究组
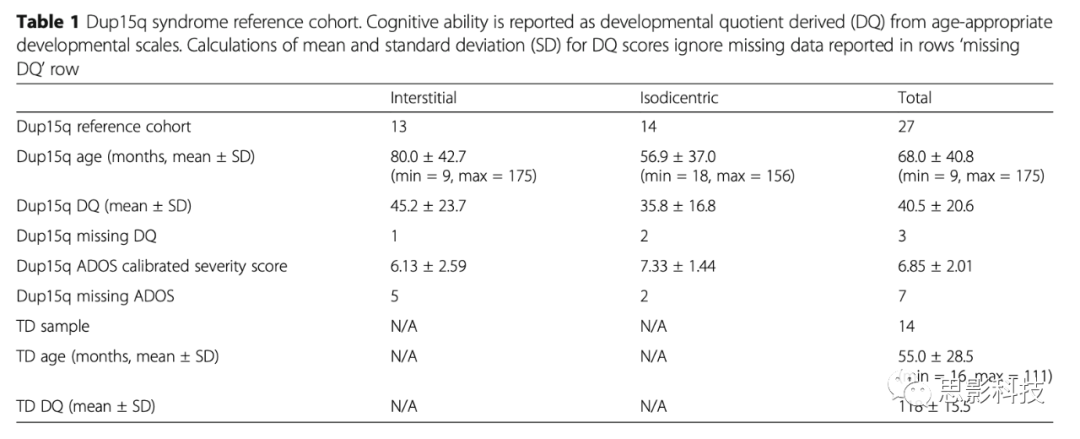
首先比较了来自TD和Dup15q研究组的所有电极和被试的平均PSD。Dup15q研究组出现显著的组水平峰值β(峰值频率:23.1±0.406Hz,平均值±标准误)。研究组峰值β功率在不同数据收集地点之间没有差异(p=0.68)。然后,比较了Dup15q研究组和TD的所有频率的脑电功率。发现相对于TD儿童,Dup15q研究组的20.2-28.5Hz功率升高(Benjamini-Hochberg p<0.05校正)。通过更大样本量证实了先前研究中报道的Dup15q综合征中存在β-功率升高现象。与TD(图1c)相比,Dup15q综合征所有电极(图1b)β活动都较高(效应大小:d′=1.06±0.325,平均值±SD),最大效应量(d>1)位于额中区(图1d)。
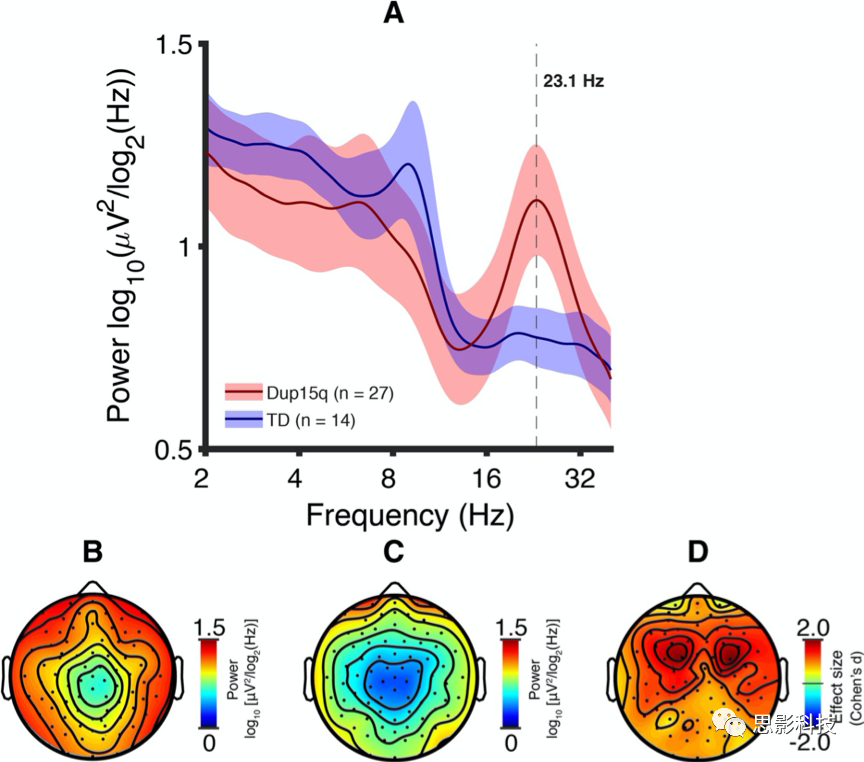
图1 Dup15q综合征与TD。
A. Dup15q研究组(红)和TD(蓝)的光谱特征。所有电极和被试的平均PSD;彩色区域表示95%的置信区间。Dup15q研究组的20.2-28.5Hz功率明显增高(p<0.05)。
B. Dup15q研究组功率地形图(23.1Hz被试平均值)。
C. TD功率地形图(23.1Hz被试平均值)。
D. Dup15q研究组与TD 在23.1Hz功率差值效应大小(Cohen's d)。跨电极平均效应量,d=1.06(最小d=0.339;最大d=1.98)
为了测试Dup15q综合征中基因数量效应,分别评估间质和等双中心倍增被试的电极平均PSD(图2a)。发现Dup15q研究组峰值β(23.1Hz)功率在倍增类型之间没有显著差异(p=0.25)。即使扩展到所有频率单元且多频率单元校正前也没有发现显著的功率差异,不过间质性Dup15q综合征在大多数频率单元有更大方差。两种倍增类型都有显著的组水平峰值β(间质峰值频率23.1±0.464 Hz;等双中心峰值频率23.2±0.567 Hz,平均值±标准误)。还观察到两种倍增类型23.1Hz的地形图相似(图2b,c)且两者功率差异效应量较小(图2d)。
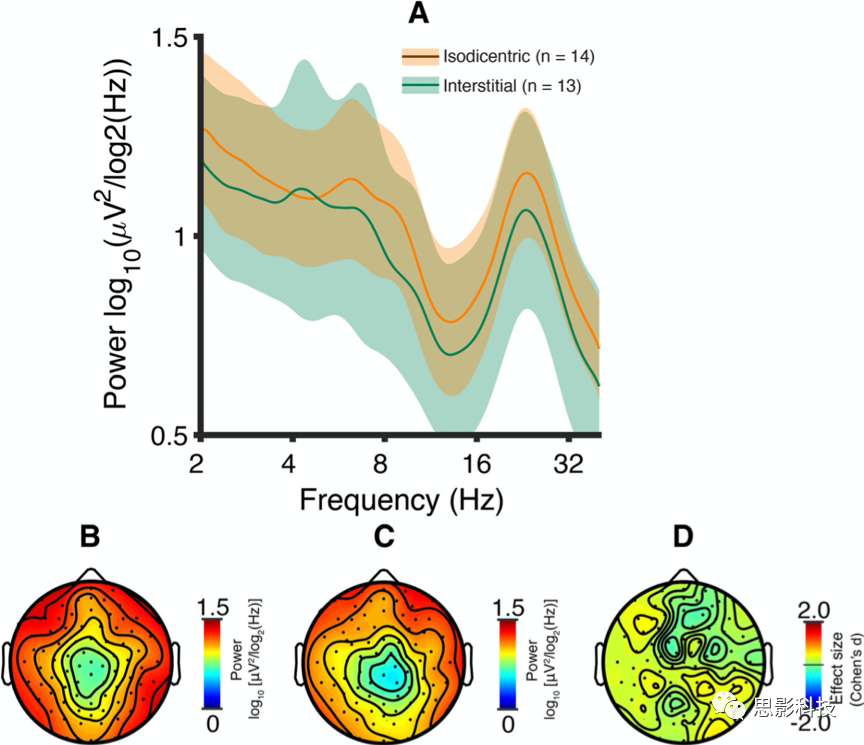
图2倍增Dup15q综合征。
A. 等双中心(橙色)和间质(绿色)倍增的光谱特征。所有电极和被试平均PSD;彩色区域表示95%置信区间。两种倍增类型在β频率都显示出显著的光谱峰(平均值:等双中心,23.2Hz;间质,23.1Hz)。倍增类型之间的功率没有差异(p>0.05,所有频率)。
B. 等双中心倍增被试23.1Hz平均功率地形图(比如Dup15q综合征插值的频率峰值)。
C. 间质倍增被试23.1Hz平均功率地形图。
D. 等双中心与间质倍增的23.1Hz功率差异效应量(Cohen's d)。跨电极平均效应d=0.21(最小d=-0.19;最大d=0.50)
β-EEG活动在Dup15q研究组和GABAA药物调节条件下类似
Dup15q研究组与GABAA-PAM(咪达唑仑,5mg口服)健康成人被试的脑电信号进行比较。在咪唑安定给药前后,α和β出现谱密度峰值(图3a)。相比基线,在Dup15q研究组峰值频率上,咪达唑仑诱发出健康被试中央区域最高β活动(23.1Hz,图3b-d)。大多数电极显示β功率增强,最大增强发生在Cz(增加76%,图4a,b)。然后,在Dup15q研究组峰值频率下对所有电极进行配对样本t检验。FDR矫正后,Fz、Cz和Pz三个电极功率显著增加(图4c)。这些电极平均功率变化的峰值出现在23.0±1.61Hz频率(平均值±标准误,增加30%,图4d),非常接近Dup15q研究组峰值频率(23.1±0.406)。
图3基线和咪达唑仑药物给药(口服5毫克)1小时后的脑电图。
A. 基线(黑色)和给药后1小时(紫色)条件下被试平均PSDs。彩色区域表示平均值95%置信区间。两种情况下,α和β频率均出现谱密度峰;咪达唑仑使α峰减弱,β峰增强。
B. 给药1h后,在Dup15q研究组峰值频率(23.1Hz)的功率地形图。
C. 基线条件下,在Dup15q研究组峰值频率(23.1Hz)的功率地形图。
D.咪达唑仑诱发的23.1Hz功率变化的效应量(Cohen's d)。最大功率变化发生在中线电极Fz、Cz和Pz(参见图3c)
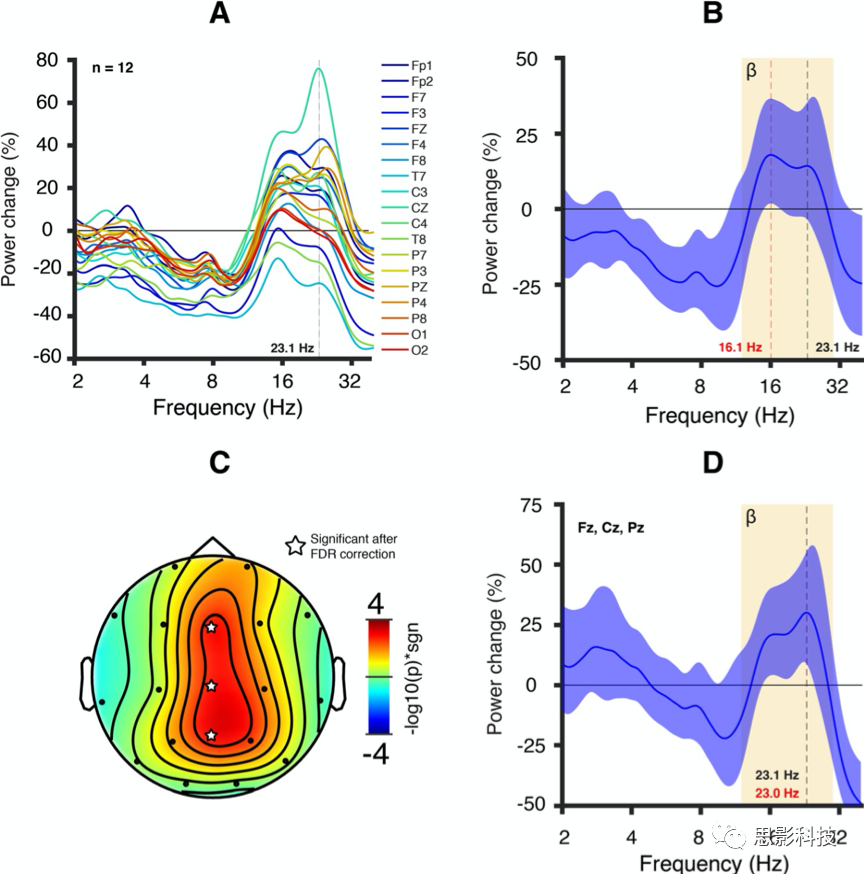
图4 GABAA受体药物调节后的健康成人脑电图特征。
12名健康成人口服GABAA-PAM(5mg咪达唑仑)。
A. 给药1小时后所有电极平均功率相对于基线条件的变化(被试绝对功率平均值)。大多数电极显示了β功率增加。
B. 电极平均功率变化。彩色区域表示95%的置信区间。平均功率变化在峰值频率(16.1Hz,红色垂直线)和Dup15q研究组峰值频率(23.1Hz,黑色垂直线)之间基本上趋于平稳。
C. -log10(p值)乘以23.1Hz双尾t检验后t值符号的地形图。多电极FDR(p =3×10-3)校正后,三个中线电极(Fz、Cz和Pz,星形符号)仍然显著。
D.中线电极功率变化平均值。经过FDR校正后23.1Hz功率变化仍然显著的中线电极功率变化。彩色区域表示95%的置信区间。功率变化峰值出现在23.0Hz(红色垂直线),非常接近Dup15q综合征的峰值频率(23.1Hz,黑色垂直线;见图1a)。
父系Dup15q综合征β-EEG活动
在两例父系Dup15q综合征案例中发现定量的β-EEG活动。这一发现在早期研究中得到了定性的观察。
1. 父系Dup15q被试801-005
801-005儿童所有PSD在β频率出现峰值(图5a,峰值频率:19.8±0.435Hz,平均值±标准误)。然后,计算了Dup15q研究组中电极平均PSD。由于研究组的年龄范围很广,使用简单的线性回归模型来解释年龄差异。父系倍增的β功率高于Dup15q研究组所有β频率平均值的95%置信区间(图5b)。801-005儿童的β功率(研究组峰值频率)位于研究组分布的上端附近(图5c,23.3Hz)。接下来,检查了研究组峰值频率的功率(重构功率)地形分布。与研究组平均地形图(图5h)相比,801-005儿童的地形图显示所有电极(图5g)有更高的功率。在所有情况下,额叶电极上出现最高功率,这也与以前的发现一致。
2. 父系Dup15q被试801-015
在801-015中观察到所有电极β频段PSD升高(图5d,峰值频率:19.3±0.677Hz,平均值±标准误)和β段内多个峰值频率(这里报告了最大峰值的频率)。这些因素会导致平均多个电极PSD后峰值不清晰,使其β频率的峰值不如801-005显著。因此本例父系Dup15q综合征与801-015儿童表现不同,但仍表现出β-EEG活动。此外,所有电极在4-8Hz(峰值频率:5.29±0.00275 Hz,平均值±标准误)都有非常明显的θ峰。 801-015儿童13.8~19.5Hz功率高于Dup15q研究组95%置信区间(图5e),19.5~27.9Hz功率在置信区间内,略低于研究组峰值频率的平均功率(图5f)。该被试在19.32+/-0.676Hz(平均值±标准误)出现峰值β,与801-005儿童的峰值频率(19.82+/-0.435Hz)和研究组峰值频率(23.1±0.406)相似。
801-015儿童 23.1Hz功率地形图(图5i),与相同频率下的研究组平均地形图(图5j)相似。801-015儿童在研究组峰值频率上双侧额叶(F3和F4)的功率最大(图5i);这与801-005的地形图相似(图5g)。父系Dup15q综合征患者中β功率升高,强烈说明UBE3A的过度表达对β-EEG活动不是必要的。
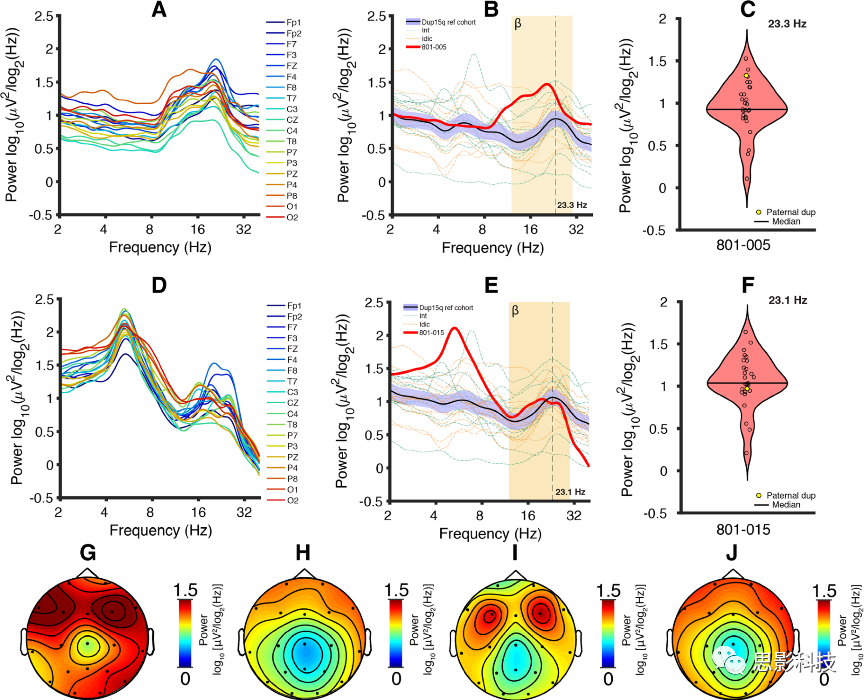
图5父系Dup15q综合征PSDs与其地形图。
父系Dup15q综合征的峰值β与Dup15q研究组的峰值β相似
为了研究母系和父系Dup15q综合征β振荡的相似性,研究了Dup15q研究组(间质和等双中心)和父系Dup15q综合征的FBPF。在Dup15q研究组中,确定了27名被试中26名的峰值β(96.3%,FBPF 22.4±2.99Hz,平均值±标准差,图6a)。研究组中,FBPF与年龄没有显著关系(p=0.25,图6b)。因此,没有用回归建模将Dup15q研究组的年龄投射到父系Dup15q综合征被试的年龄上。在研究组中,倍增类型(间质型和等中心型)之间的FBPF没有显著差异(p=0.085)。在父系Dup15q综合征中,两名被试的FBPF均在研究组平均值的一个标准差范围内(图6c;801-005:FBPF 20.7Hz,z=-0.59;801-015:FBPF 20.1Hz,z=-0.78 Hz)。父系Dup15q综合征被试在频率-功率分布上与Dup15q研究组聚合在一起(图6a)。
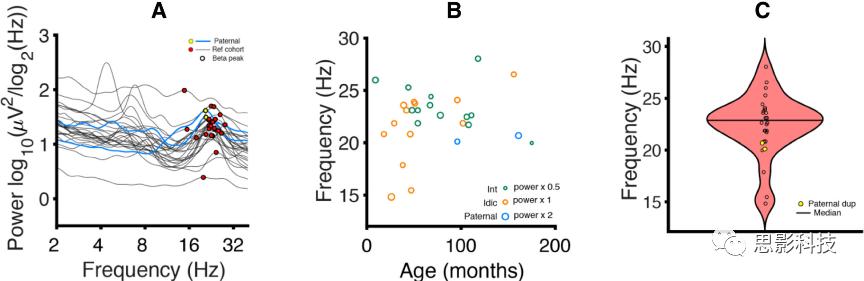
图6峰值频率分析。
A. 来自Dup15q研究组(黑色)和父系Dup15q综合征(蓝色)的PSDs。β峰标记为红色(研究组)和黄色(父系Dup15q)。两个父系Dup15q综合征的β峰似乎都属于研究组中发现的β峰群。
B. 年龄与FBPF。点代表被试,点大小代表相应的FBPF的峰值功率(绿色=间质研究组,橙色=等中心研究组,蓝色=间质父系Dup15q综合征)。年龄与FBPF无关(p=0.25)。
C小提琴图。黄点代表父系Dup15q综合征。父系Dup15q综合征的两个被试都在研究组的一个标准差内。
结论
基于机制产生且可以指导临床试验的定量生物标记物,是神经发育障碍研究十分需要的。本研究获得了Dup15q综合征稳定脑电标记物生物机制的有价值证据。这种生物标记物不能简单用UBE3A水平升高本身来解释,但可以通过健康成人的GABAergic调节来重新解释,说明该β-EEG活动可能是Dup15q综合征中GABAA活动增加或对GABA敏感的外在表现。本研究是在分子机制中建立Dup15q综合征生物标记物并促进其在临床试验应用的重要一步。

微信扫码或者长按选择识别关注思影
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
更新通知:第十届脑影像机器学习班(已确定)
磁共振脑影像结构班(预报名)
弥散磁共振成像数据处理提高班(预报名)
小动物磁共振脑影像数据处理班(预报名)
更新通知:第二十届脑电数据处理中级班(已确定)
脑电信号数据处理提高班(预报名)
眼动数据处理班(预报名)
近红外脑功能数据处理班(预报名)
数据处理业务介绍:
招聘及产品:
招聘:脑影像数据处理工程师(重庆&南京)