

睡眠对于认知和维持健康的大脑功能至关重要。神经活动中的慢波有助于记忆巩固,而脑脊液(CSF)有助于清除大脑中的代谢废物。这两个过程是否相关尚不清楚。波士顿大学生物医学工程系的Fultz等人对此进行了研究,结果发表在Science杂志。我们使用累加的神经影像技术来测量人脑的生理和神经动力学。发现非快速眼动睡眠期间出现的振荡电生理,血液动力学和CSF动态的连贯模式。神经慢波之后是血液动力学振荡,而血液动力学振荡又与CSF流量相关。这些结果表明,沉睡的大脑在宏观范围内表现出CSF流动波,并且这些CSF动态与神经和血液动力学节律相互关联。

具体研究:
睡眠对于高级认知加工以及生理功能的基本维持和恢复都是至关重要的。在人类非快速眼动(NREM)睡眠期间,脑电图(EEG)表现出低频(<4 Hz)振荡动力学,可支持记忆和神经计算。此外,功能磁共振成像(fMRI)研究测量了血氧水平依赖性(BOLD)信号,已证明 NREM睡眠期间广泛的血液动力学改变。睡眠还与组织液量增加和代谢废物进入脑脊液的清除率相关,并且睡眠中的清除率随着低频脑电图振荡的增加而增强。在这种低唤醒状态下为什么会同时发生这些多样的生理过程,这一点尚不清楚。特别是,脑脊液动力学在睡眠中是如何变化的,以及它们与神经活动和血流动力学的主要变化有何关系,目前仍不清楚。
我们同时测量了人类睡眠过程中的BOLD的动态,EEG和CSF流量。为了获得高时间分辨率的成像,我们以快速的速率[重复时间(TR)<400ms]采集了fMRI数据。尽管功能磁共振成像通常用于检测局部氧合变化,但快速采集范例还可以检测流体流入:到达成像空间边缘的新鲜流体信号强度很高,因为它尚未经历过射频脉冲。通过将成像体积的边界放在在第四脑室(图1A)中, CSF流入大脑的信号在下部切片中被检测为增加的信号(图1B),这使我们能够同时测量BOLD与CSF流动的动态。我们将这种影像学与同时进行的脑电图(n =13 名被试)相结合,并识别出清晰稳定的唤醒或NREM睡眠,从而能够分析连续的低频动态。
我们首先研究了睡眠是否与独特的脑脊液流动动力学有关(图1,C至E)。在觉醒期间,CSF信号在〜0.25Hz表现出与呼吸信号同步的小振幅节律(图1,E和G),与先前的研究一致。相比之下,在NREM睡眠期间,我们观察到CSF信号在0.05Hz时出现了较大的振荡(图1,E和G)。我们分析了所有睡眠节段的CSF信号,确认已识别的睡眠节段表现出NREM睡眠的低频EEG特征(图1F)。我们发现睡眠期间CSF信号在0.05Hz峰值处增加了5.52dB(图1,G和H)[95%置信区间(CI)=(2.33,7.67);P=0.003],表明大约每20s就会发生一次CSF流入的大的波动。我们还分析了附近的非CSF感兴趣区域(ROI),没有发现这种影响[change=-0.03dB,CI=(-2.7,1.3)],表明这种睡眠相关模式是由心室中的生理信号特别驱动的(图1I)。由于流入信号是由流入采集空间的流体引起的,因此CSF流量信号在边缘中应最亮,并在流体进入中心时衰减(图2,A和B)。我们确实观察到了整个切片上信号幅度的梯度(图2,C和D)。一些较大的流入事件在较低的切片上表现出同样明亮的振幅(图2D),表明脑脊液流速超过成像临界速度。这些结果确定了在NREM睡眠期间出现的0.05Hz的CSF大幅度流动。
接下来,我们检查了这些缓慢的宏观CSF振荡是否与血液动力学信号有关。先前的研究提出了对应于〜1Hz心跳周期的微观动脉搏动,作为驱动组织液流动的一种机制。为了测试我们观察到的可能导致慢得多的宏观CSF节律,我们分析了睡眠期间BOLD信号的变化。我们观察到,与清醒相比,睡眠期间灰质的BOLD信号幅度增加(图3A与B和C)[平均值=3.28dB;CI=(0.09,6.54);P=0.032],与以前的睡眠期间低频BOLD波动的报告一致。此外,CSF信号在睡眠过程中在时间上紧紧耦合到皮层灰质BOLD振荡(图3,A和B),表现出很强的反相关性(在滞后2s时最大r=-0.48,P<0.001)。
这种反相关性提示睡眠期间血液流量和CSF流量可能会发生变化。我们假设,BOLD振荡对应于脑部血容量的振荡,并且由于颅内容积恒定,当血液占据的容积较小时,更多的CSF流入头部。该假设预测,在将负值设置为零之后,CSF信号应大致匹配BOLD振荡的负导数。与此假设一致,CSF时间序列与阈值导数BOLD信号高度相关(图3,D和E)(滞后-1.8s时最大r=0.59;P<0.001)。
接下来,我们检查了神经活动是否与睡眠期间这些耦合的血液动力学和CSF振荡有关。在常规功能磁共振成像中,bold反应是由神经活动引起的,神经活动驱动了富氧血液的流动,而脑电图的振荡与血液动力学信号有关。因此,我们假设NREM睡眠的大振幅的慢delta波(0.2-4Hz)的电生理振荡可能与血容量的振荡有关,进而对CSF流量产生位移影响。我们分析了与CSF波峰值有关的慢delta的瞬时振幅,发现神经波,BOLD波和CSF波是耦合的(图4,A至C)。神经波先于CSF波,慢delta波的峰值在CSF峰值之前6.4s出现(峰值幅度=21%,P<0.001)。我们计算了脑电图和脑脊液之间的最佳拟合脉冲响应(图4D)并且发现将脑电图与该脉冲响应进行卷积可以显著预测脑脊液动力学(zero-lag r=0.23,交叉验证r=0.22±0.07)。为了检查这种EEG-CSF耦合是否与睡眠中CSF波的出现特别相关,我们测试了EEG对CSF波最大的部分中CSF动态的预测情况。脑脊液波较大的部分的表现较高(zero-lag r =0.54, P < 0.001),表明在CSF最丰富的睡眠段中,EEG与CSF的联系更紧密。

图1.睡眠期间,第四脑室出现CSF信号的大振荡。
(A)扫描定位示例。黄粗线:位置相对于解剖结构的功能图像。底部边缘与第四脑室相交(红色箭头),可以测量CSF流入量。
(B)来自底部切片的示例功能像。通过心室的流入被检测为亮信号(红色箭头)。
(C)脑电图显示了长时间的NREM睡眠和唤醒(枕叶 α -10Hz)。
(D)行为反应。
(E)单个CSF体素的时间序列(使用10-TR的平滑核)显示出较大的,缓慢的睡眠动态,在清醒期间逐渐消退。
(F)枕叶EEG的平均功率谱密度(PSD)证实了睡眠中的慢delta功率,而不是清醒的高α功率。
(G)CSF信号的PSD显示睡眠期间的功率为0.05Hz增强(n=13位参与者睡眠;11位参与者唤醒)。阴影区域表示95%的置信区间。
(H)在睡眠期间的低频(LF,0至0.1Hz)CSF功率增加(成对比较,n=11位参与者)。
(I)这种睡眠选择功率增加是特定于心室ROI的,在相邻大小匹配的对照ROI(n=11位参与者)中未观察到。

图2.对应于〜0.05Hz的脑脊液搏动的心室信号。
(A)流入大脑的新CSF将产生明亮的信号。
(B)流入信号在底部切片中最大,并且向内振幅减小。如果流量超过临界速度,则底部的CSF切片被完全替换,并且内部切片中的信号幅度也很大。
(C)切片的平均幅度在切片数增多时衰减。
(D)第四脑室成像体积底部切片的时间序列在较低的切片(如第2脑室)显示的信号最大,在较高的切片(如第4脑室)显示的信号较小。橙色箭头表示流速(较大的箭头表示较高的流速),黑色箭头表示单个事件。
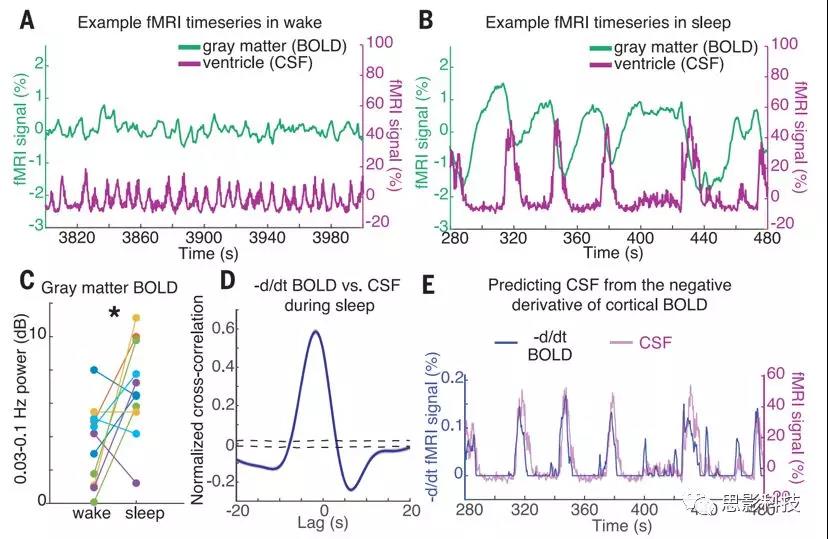
图3.CSF振荡与睡眠期间出现的皮质灰质中的血流动力学振荡反相关,当血液量减少时,CSF流量增加。
(A)一名参与者的皮质灰质BOLD信号和平均CSF信号的时间序列示例。在唤醒过程中,信号为低振幅并与呼吸同步(0.25Hz)。
(B)在睡眠期间,出现大幅度的BOLD振荡,并且其时程与心室CSF信号(〜0.05Hz)耦合。
(C)睡眠期间平均皮质灰质BOLD信号功率增加(n=11个参与者,配对测试)。
(D)BOLD的零阈值负导数与CSF信号之间的平均互相关显示出很强的相关性(n = 176个片段,13个参与者)。蓝色阴影区域表示分段的标准误差;黑色虚线表示打乱分布的95%置信区间。
(E)时间序列显示出相关性,表明当脑血容量减少时CSF向上流至第四脑室。
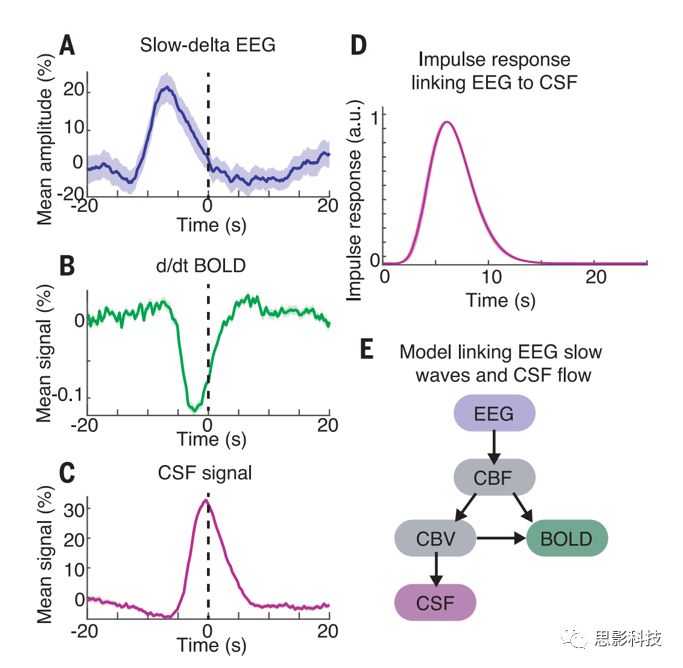
图4.脑电图慢波delta与bold信号和脑脊液的波是耦合的。
(A)慢速delta的平均波幅包络线;
(B)BOLD信号的平均导数;
(C)平均CSF信号,所有这些信号在睡眠期间都锁定在CSF波的峰值上;
(D)CSF信号对EEG包络的脉冲响应显示出与以前建立的血液动力学模型相似的时间过程;
(E)将神经活动的时间过程与CSF流量联系起来的模型图。变量包括CBF和脑血容量(CBV)。
因此,EEG,BOLD和CSF信号的相干动力学表现出特定的时序,在随后的BOLD和CSF波之前具有神经节律。为了测试这些观察到的相关性和延迟模式是否可能源于生物物理机制,我们使用已建立的血液动力学耦合模型构建了一个计算模型(图4E)。我们提取了delta波的包络线,由于其对神经活动的抑制作用,预期与脑血流量(CBF)信号的减少相关。然后使用神经振荡来预测血流,血容量和脑脊液的时程。该模型首先使用先前报告的生理参数,没有其他参数拟合,以测试我们的观察是否与这些信号之间已建立的生物物理耦合一致。模型预测结果显著大于所有滞后项下原始脑电图包膜与脑脊液的最大相关系数。该模型的预测显着大于所有滞后的原始EEG包络线与CSF之间的最大相关性(最大r=0.15,P<0.05),表明我们的数据与神经活动,血液动力学和CSF之间的生物物理耦合一致。
我们得出的结论是,人类睡眠与大的低频神经元活动,血液氧合和CSF流量的耦合有关。尽管众所周知,电生理慢波在认知中起着重要作用,但我们的结果表明,它们也可能与睡眠的生理恢复作用有关,因为慢的神经活动会伴随着全脑的血容量和脑脊液流量的搏动。
这些结果解决了睡眠神经生理学中一个关键的缺失环节。我们发现的CSF流动的宏观变化预计会改变废物清除,因为脉动流体动力学可以增加混合和扩散。已经提出的神经血管耦合有助于清除废物,但是为何在睡眠期间会导致较高的清除率尚不清楚。我们的研究表明神经和血液动力学缓慢振荡与其他生理因素相结合,可能是造成这一过程的原因。未来的动物研究可以测试这些神经和生理节律之间的因果关系。
我们对与睡眠相关的CSF流体动力学的研究还表明,在与睡眠障碍相关的临床疾病中将探索潜在的生物标志物。衰老中的记忆障碍与慢波的抑制有关;我们的模型表明,这种慢波损耗将与CSF流量减少有关。此外,我们的结果暗示了最近发现之间的潜在联系,即tau CSF水平和依赖于睡眠和神经活动的淀粉样β,而振荡性神经活动导致tau减少,相干神经活动可能表示更高的蛋白质清除。综上所述,我们的研究结果确定了睡眠中出现的CSF流动,并表明神经活动的缓慢节律与这些CSF波相互关联,血液动力学振荡是这些过程相互耦合的中间机制。

微信扫码或者长按选择识别关注思影
第十四届磁共振脑网络数据处理班(重庆,7.26-31)
第三十届磁共振脑影像基础班(南京,7.31-8.5)
第十届脑影像机器学习班(南京,6.30-7.5)
第十二届磁共振弥散张量成像数据处理班(南京,6.18-23)
第二十届脑电数据处理中级班(重庆,8.9-14)