

德国海德堡大学的Adriano B.L. Tort,Jurij Brankack和Andreas DraguhnTrends in Neurosciences最近发文。作者回顾了最近的研究,发现啮齿动物的一些脑区内,鼻腔呼吸诱发的神经震荡频率与呼吸频率相同。此外,呼吸可以调节特定γ子频段(70–120Hz)的振幅,尤其在额叶最为显著。由于啮齿动物通常以δ和θ频率呼吸,因此作者特别关注了δ和θ功率及两者跨区域同步性,以及δ-γ和θ-γ耦合的已有研究,以期探究呼吸诱发节律与呼吸-γ耦合。作者认为同时记录呼吸和电生理数据对正确识别大脑振荡是必要的,并假设呼吸诱发的振荡有助于大脑中的远程信息传递。
呼吸诱发震荡是一种全脑节律
自Lord Adrian在20世纪40和50年代的开创性工作以来,鼻呼吸会使神经振荡(neuronal oscillations)频率锁定在嗅球(OB)呼吸周期左右就已众所周知。在21世纪,Fontanini和他的同事已经证明呼吸诱发振荡(respiration-entrained
oscillations)会传播到初级嗅觉(梨状)皮层(图1A)。最近,人们越来越认识到呼吸也会影响一些更深层的下游脑区的网络激活。例如,在清醒且静止小鼠的胡须体觉皮层(whisker barrel cortex)的局部场电位(LFP)中已观察到呼吸驱动的δ频率范围(0.5-4Hz)内的震荡(图1B)。呼吸诱发的LFP振荡也出现在大鼠和小鼠的背海马区,麻醉或清醒状态下最显著的区域均在齿状回(DG)。其瞬时频率在δ或θ(4-12Hz)范围内,具体取决于呼吸频率(图1C)。值得注意的是,最近有研究报告在静止和活动行为中内侧前额叶皮层(m PFC)均具有显著的呼吸诱发振荡(图1D)。其他研究也扩展了这些发现,并表明呼吸锁定的这类激活也发生在人类大脑的几个脑区。
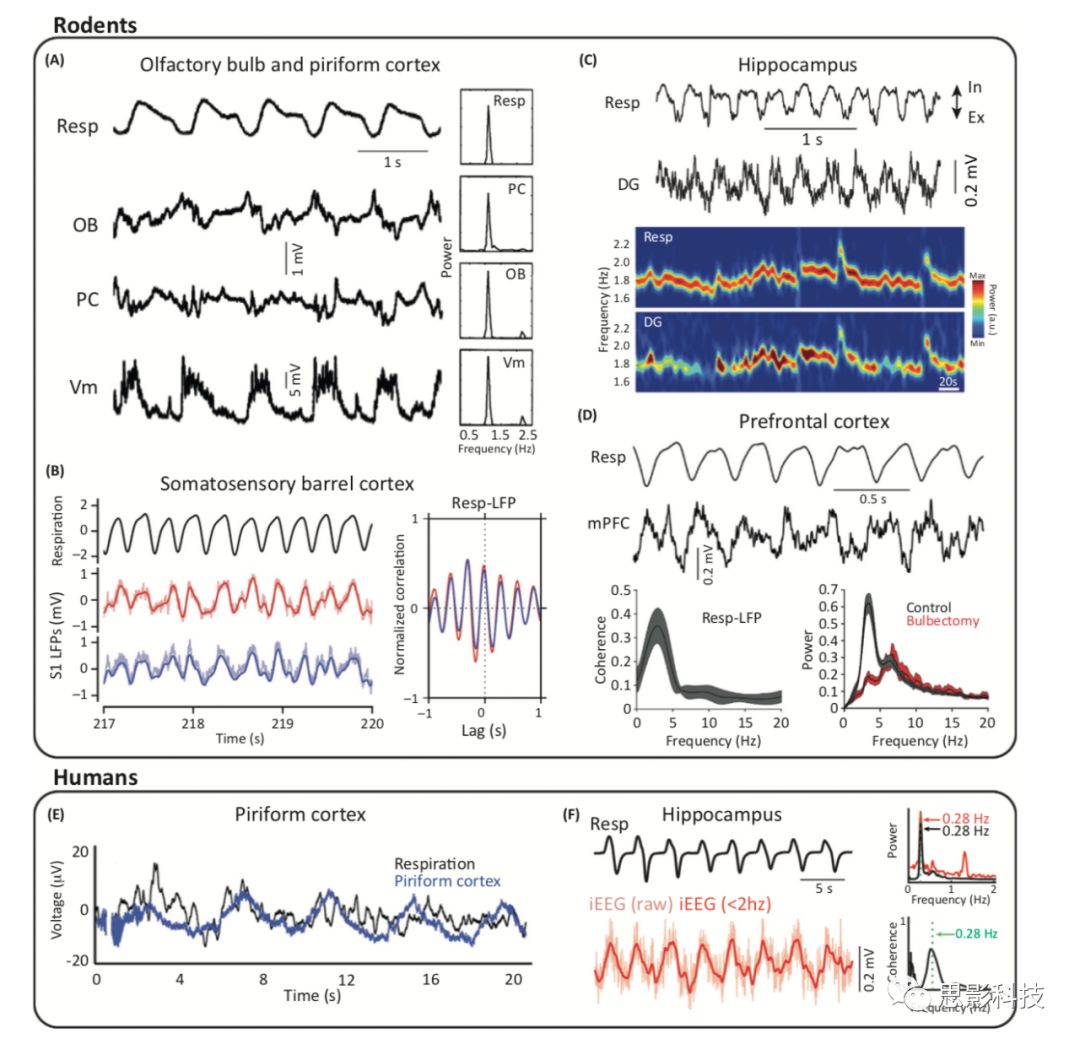
图1. 广泛脑区和跨物种的呼吸诱发局部场电位震荡。
(A)左侧表示呼吸(Resp),及同时记录于嗅球(OB)和梨状皮层(PC)的LFP,以及PC神经元(Vm)的细胞内记录,数据取自氯胺酮麻醉的大鼠。右侧表示相应的功率谱。
(B)左侧表示呼吸(黑色)和两个LFP(红色和蓝色),三者同时记录于清醒且头部固定的小鼠须状桶皮层。右侧表示LFPs与呼吸之间的交叉相关。
(C)上侧表示呼吸和同时记录的来自氨基甲酸乙酯麻醉鼠齿状回(DG)的LFP。下侧表示呼吸频率和峰值频率相应的变化。
(D)上侧表示清醒静止小鼠的呼吸频率和内侧前额叶皮层的LFP。下侧左部分表示LFP与呼吸的高度一致性,下侧右部分表示呼吸的频率随着嗅球频率变化而消失。
(E)顽固性癫痫患者同时记录的呼吸(黑色)和PC LFP(蓝色)。
(F)左侧表示癫痫患者海马内的呼吸节律(黑线)和颅内脑电图(iEEG,红线)。右侧表示呼吸和海马信号的功率谱(顶部),以及它们的相干谱(下)。
当前有大量的证据表明,呼吸诱发的振荡并不是由呼吸相关的肌肉活动或电极移动等伪迹所致。例如,它们可以调节动作电位和sharp-wave ripples的概率;它们在嗅球切除后或当动物通过气管而不是鼻子呼吸时会消失;在海马区,它们表现出清晰的层次轮廓,并在DG中振幅最大。重要的是,尽管存在频率重叠,这些振荡可以通过多种特征从海马产生的θ波中区分,如通过最大振幅层、呼吸一致性、对鼻息的依赖以及对胆碱能传递不敏感等。同样,呼吸诱发节律也可从与丘脑激活相关或与睡眠和深度麻醉期间up-and-down状态转换相关的δ频率中区分出来。
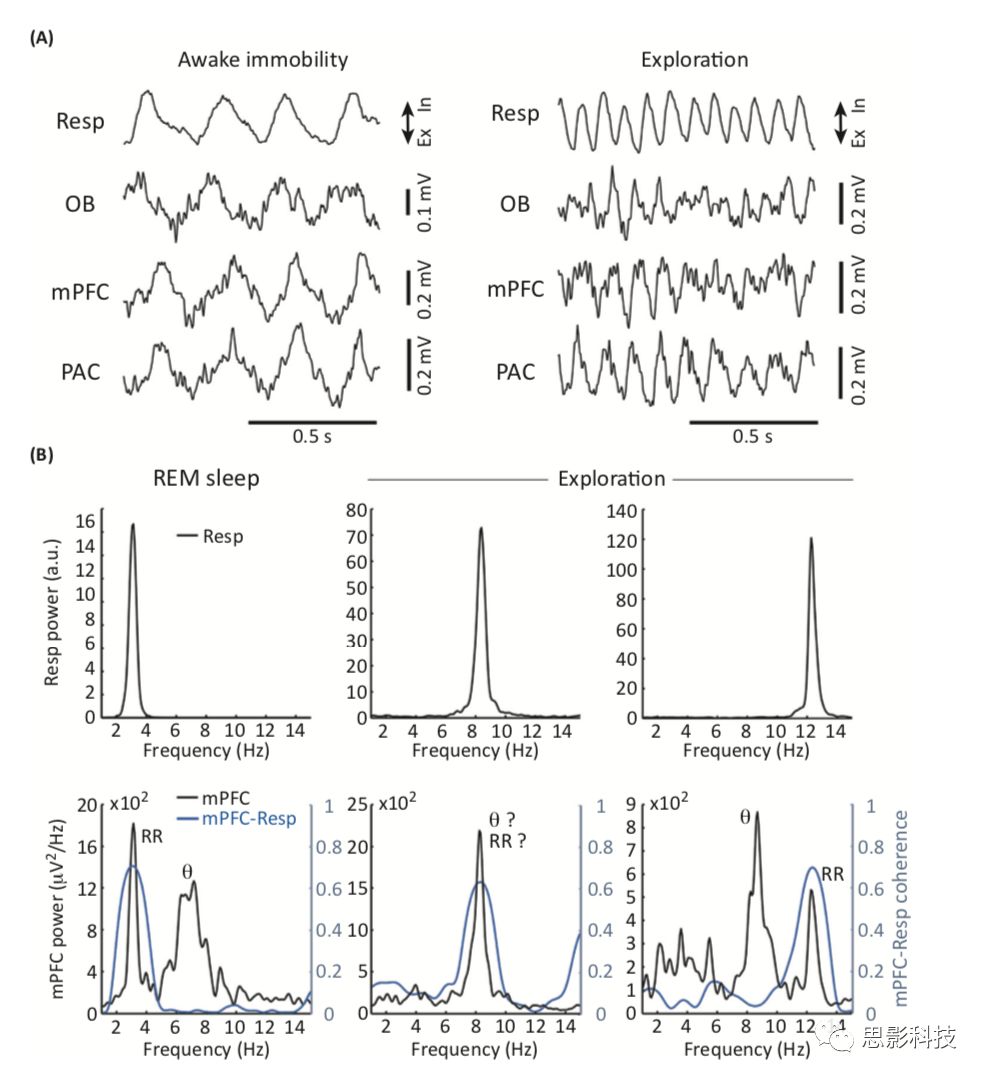
图2.呼吸诱发LFP震荡与θ震荡的共存。
图2A展示了清醒静止小鼠自发呼吸所诱发的LFP振荡,同时记录有小鼠的OB、mPFC和顶叶皮层的节律变化。需要注意的是,在静止状态下,呼吸锁定的振荡发生在δ频率,并且可以很容易地从所有记录脑区(呼吸频率=~3.8Hz)的LFP轨迹中推断出来。然而,在探测过程中,由于呼吸频率较快而且伴随着θ波的存在(θ频率=~9.2Hz;呼吸频率= ~11.5 Hz),它们很难通过视觉检测发现,尤其是神经皮层中的LFPs更难区分。在这种情况下,只有通过分析LFP功率谱(在附近频率上显示出两个峰值),并通过分析LFP与呼吸的相关性才能确定这两个振荡,以表明哪一个峰值是与呼吸周期相位锁定的。
图2B展示了呼吸频率不同的三个时期内的鼻呼吸(上)和mPFC的LFP信号(下)的功率谱,以及LFP和鼻呼吸之间的相干谱。图2A中的探索示例类似于图2B中最右边的情况。可以在功率谱中看到θ的振荡(多数情况出现于快速眼动阶段),但是,与呼吸诱发节律不同的是,θ波与鼻呼吸并不存在相干性。此外,随着呼吸速率的变化,呼吸诱发LFP节律的频率也会发生变化,而θ节律的峰值频率则要稳定得多。重要的是,啮齿类动物的呼吸频率可以从休息和睡眠中的~1Hz到嗅时的~14Hz之间变化。因此,呼吸锁定节律的频率可以比θ频率慢、相类似或快(图2B)。这有着重要的意义:作者认为以前关于θ频率振荡的研究可能实际上检测到的是锁定于呼吸的LFP相位活动,而不是被定义在septohippocampal回路中的经典θ节律。尽管在自由活动的动物中,CA1 θ的峰值频率通常在6~9Hz之间,但大多数研究人员将θ振荡定义在4~12 Hz之间,这很可能增加呼吸诱发节律产生“污染”的几率。值得注意的是,Macrides等人的开创性工作,其中Kay报告了认知任务中海马和OB在θ频率上的同步。但是,这种同步性是发生在经典的θ节律上,还是发生在呼吸诱发振荡后的θ频率上还有待确定。在当时,呼吸诱发振荡并不被认为是一种独立于海马中θ的活动。
作者注意到,类似的问题可能发生在关于δ频率振荡的研究中,特别是在较大的啮齿类动物中,如大鼠,它们的呼吸速度比小鼠慢。例如,大鼠在气味工作记忆任务中,所谓的与mPFC、腹侧被盖区和海马相关的4Hz的振荡可能对应着呼吸诱发振荡。最近,麻醉大鼠mPFC中2-5Hz振荡的起源也被认为是呼吸引起的。有趣的是,最近的一项研究表明,在社会互动过程中,草原田鼠的mPFC和伏隔核的振荡频率相似,但其起源尚未确定。作者还认为,在恐惧条件化中小鼠出现冻结行为时,mPFC和杏仁核中出现的4Hz振荡是由呼吸引起的。相比之下,睡眠相关的慢波振荡先前被认为是呼吸引起的,而最近的研究发现了两者的相对独立性,尽管它们的频率有所重叠。
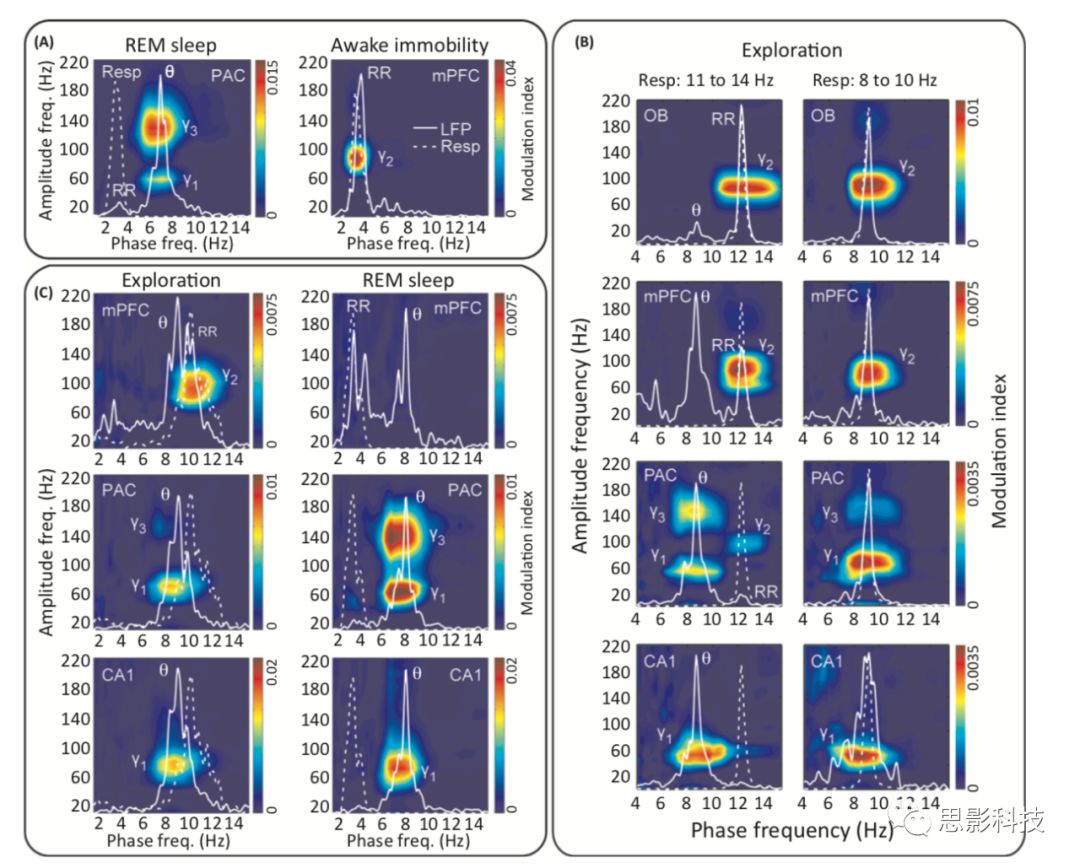
图3. θ与呼吸调节不同的γ频率。
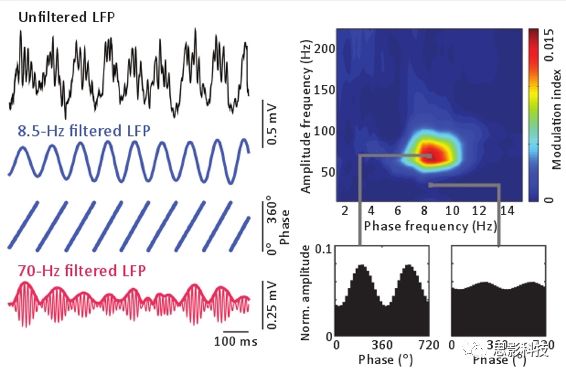
图4.通过Comodulation Maps评估相位-幅值耦合。
θ和呼吸调节不同的γ频率
对θ-γ耦合的研究表明,大范围的γ频率(30~160Hz)可分为独立的子频带。随着现代信号分析工具的发展,如comodulation map的出现,使得这种划分可以实现(图4)。在啮齿类动物的海马和顶叶皮层,comodulation map显示,θ相位在大范围的γ频率内调节多个不重叠的振荡子频带。此外,不同的γ子频带发生在不同的θ相位和θ周期上。因此,相位-振幅耦合的分析已经开始揭示一种先前没有被功率谱分析所揭示的γ振荡的更细化的功能划分。此外,这些研究还表明,跨频耦合模式取决于沿顶叶-CA1-齿状轴记录的层数,以及诱发θ的行为类型(例如,运动与REM睡眠)。图3A(左)显示的是在REM睡眠期间从小鼠顶叶皮层记录的LFP的comodulation map。注意,θ相位有力地调节了两个不重叠的γ子频带的振幅:~40-90-Hz和~110-160-Hz振荡。
呼吸周期的相位一直被认为可以调节OB中的γ振幅。此外,Fontanini和Bower阐述了呼吸周期在梨状皮层中引发γ活动激增。最近,也发现呼吸调节非嗅觉区内的局部γ。即2014年Ito等人证明呼吸诱发振荡(δ频率)对静止小鼠胡须感觉皮层中的γ有调节作用,而呼吸-γ耦合则在麻醉小鼠的海马内被发现。2017年,Biskamp等人以及Zhong钟等人发现,清醒小鼠的mPFC表现出明显的呼吸振荡和呼吸-γ耦合。值得注意的是,根据呼吸频率的不同,comodulation map中的呼吸-γ耦合可以解释为“δ”-γ耦合(在慢速呼吸期间)或“θ”-伽玛耦合(在快速呼吸期间)。
Comodulation
maps还揭示了呼吸调节着一个特定的伽马子频带,这与θ调制的γ子频带是不同的(图3A)。奇怪的是,呼吸调节的γ(70~120Hz)的频率范围位于θ调节的两个γ子频带之间(比较图3A的左右两侧的激活区)。但有趣的是,Kay以前曾报道过OB有两种γ活动(35~65Hz和65~100Hz),而且只有频率较快的γ子频带受到嗅探周期较强的影响。与Kay的发现一致,Zhong等人也发现了OB在40-70 Hz内的一个较慢的γ活动没有与呼吸耦合,因此在comodulation map中并不明显。总之,这些结果表明,在较宽的γ范围内不同频率通道可用于区分不同的信息类型。
前额叶皮层中γ震荡与呼吸之间的选择性耦合
γ振荡与θ或呼吸诱发LFP节律的耦合取决于记录的区域和行为状态。例如,静止期间,OB、mPFC、顶叶皮质和海马在70到120Hz振荡与呼吸之间表现出耦合。相反,在运动、吸嗅和探索等活动中,耦合可分为两种区域模式:
额叶区(OB和mPFC)与呼吸在70-120Hz的γ子频带中仍表现出专属耦合,而海马和顶叶皮层则在40-90 Hz和110-160Hz的γ子频带表现出耦合。图3B(左)显示了在探索过程中每个区域的有代表性的comodulation map,以及LFP与呼吸的功率谱。在mPFC中,γ耦合只与呼吸有关,而与θ无关。这种特异性是值得注意的,因为在这个区域的LFP同时包含了这两个慢振荡,从功率谱的两个峰值可见。Biskamp等人报告一致发现,在mPFC中观察到80-100Hz的γ明显受到呼吸诱发振荡的调节,而θ在这一区域没有表现出与γ有意义的耦合。
最近的研究表明,γ在mPFC中选择性地与呼吸相耦合,因此以往那些在前额发现θ激活以及θ-γ耦合的报告是需要重新审视的,因为他们可能探测的是呼吸锁定的节律却把它当作θ振荡。如在图3B(右)所示,其中呼吸的频率就与CA1θ节律相同。在这种情况下,频谱分析无法确切地区分θ、呼吸诱发节律以及这两种节律的叠加。由于呼吸的频率与海马内θ频率相似,人们很容易得出结论:θ在mPFC和OB中调节70-120Hz的振荡。由于以前的大多数研究都没有同时记录LFPs与呼吸的节律,所以他们的发现是否受呼吸诱发LFP节律的影响还有待确定。调节γ活动的确切频率范围可能会为确定慢波振荡的性质提供一些线索;但是,尽管θ和呼吸调节的γ子带在动物体内不同,但在某些情况下,它们可能部分或甚至基本上重叠,特别是在分析不同的动物时。除了调节γ频率的差异外,还有另一个生理特征可能有助于将θ-γ耦合与呼吸-γ耦合分离开来,这是我们接下来讨论的。
在REM睡眠期间,γ与呼吸耦合减少,而θ-γ耦合增加
在快速眼动睡眠中,θ-γ耦合一直在增加(图3C)。这种效应在110-160Hz的γ子频带中的θ调节最为明显。与之形成鲜明对比的是,呼吸-γ耦合在快速眼动睡眠中会大大减少,甚至消失,尽管LFP仍然表现出呼吸诱发节律(图3C)。有趣的是,最近的两项研究表明,在清醒状态下mPFC中观察到的θ-γ耦合在快速眼动睡眠中消失了。虽然呼吸在这些研究中没有被同时记录,但鉴于对快速眼动睡眠的负性依赖,作者怀疑在清醒状态下观察到的耦合模式实际上对应的是呼吸-γ耦合,而不是θ-γ耦合。因此,当呼吸没有被直接监测时,两个特征可用以甄别慢波振荡的性质:调节γ振荡的精确频段和耦合对快速眼动睡眠的依赖性。但需要注意的是,这种差异仅在小鼠身上有报道,而在大鼠和其他啮齿类动物中还并未发现。
呼吸诱发节律的潜在机制与功能
嗅觉区的神经网络,如OB和梨状皮层,与呼吸周期有很强的同步,这是由LFP和尖峰活动推测的。先前的工作表明,这种相位锁定依赖于鼻腔上皮细胞对感觉的再输入,而不是负责控制呼吸输出运动的脑干回路中的中央模式产生器。值得注意的是,虽然锁相于感觉传入的神经活动依赖于鼻息,但作为刺激的空气并不需要含有气味。目前的证据表明,在非嗅觉区域的呼吸诱发振荡也依赖于来自鼻腔的感觉信号,而不是呼吸中枢模式产生器。例如,通过气管切开或OB的失活阻碍体感皮层、海马和mPFC内的呼吸锁定激活(见图1D),而动物仍然表现出呼吸运动。此外,可以通过有节律的将空气喷射入鼻腔而诱发嗅觉区和非嗅觉区的神经激活。主要嗅觉区是将呼吸节律传递到下游脑区的首选脑区。如我们已熟知的解剖联系:梨状皮质与前额叶皮层的直接联系,以及梨状皮层通过内嗅皮层与海马的间接联系。然而,其他区域,如胡须体觉皮层,则可能依赖于多突触通路。
呼吸-γ耦合的机制目前尚不清楚。为了深入了解这个问题,Heck等人提出了一种通过大量短距离和少量长距离连接以激发或抑制细胞耦合的graph模型,以重新描述皮层网络的结构。呼吸效应被模拟为正弦输入模型;呼吸输入诱发扰动,使轨迹进入γ周期有限循环(吸气期间)的相位或离开该相位(呼气期间)回到定点。因此,该模型提供了一个证明准则,假设呼吸驱动神经元兴奋性,那么被呼吸周期调节的γ振幅可能是通过皮层网络的固有特性来实现的。虽然呼吸确实调节了几个脑区的尖峰概率,但后者是Heck等人在逻辑上模拟的现象,它的生物物理/突触起源仍有待确定。另一种假设是γ通过GABAergic在中间神经元的抑制性传递产生,而这些神经元将直接受到来自嗅觉区呼吸锁定输入的调节。中间神经元一贯被认为与γ存在联系,但对于不同的γ子频带与中间神经元亚型的多样性联系人们知之甚少。此外,嗅觉相关输入会冲击大脑广泛区域的中间神经元是否存在还有待证实。
有趣的是,在人类身上的一些呼吸练习,如冥想,已经被证明会影响情绪状态和认知功能。无论其潜在机制如何,鼻腔呼吸都有可能通过驱动慢波振荡和调节快波振荡的幅度来影响大脑的功能。与此一致的是,目前的证据表明大脑振荡及其跨频耦合发挥了功能性的作用。呼吸诱发的慢波振荡往往在很长的距离内是连贯的,这非常适合于脑区间的交流,而速度更快的γ振荡则与局部运算相关联。因此,呼吸-γ耦合可以作为一种跨远距离脑区整合局部γ的方法,这类似于先前提出的关于θ-γ耦合的方法。重要的是,由于θ和呼吸调节不同的γ频率,不同类型的信息可以在不受干扰的情况下同时在大脑中传播。虽然这都是假设,但至少了解呼吸对大脑的影响有助于理解大脑振荡的内在联系。
到目前为止,有关呼吸诱发节律的大多数发现都是从小鼠和大鼠身上获得的,但最近的两项研究表明,人类大脑中的电活动也锁定于呼吸周期,并与啮齿类动物一样都依赖于鼻息。这些研究还表明,呼吸周期调节了颅内脑电(EEG)高频振荡的幅值,而且不同的呼吸路径(鼻腔和口腔)影响健康被试的认知能力。值得注意的是,相位-幅值的跨频耦合在人类中已经得到了很好的证实,以前就有发现其与感觉、运动和认知功能有关。因此,更快的振荡耦合于呼吸周期这一事实与鼻呼吸可以调节高级脑功能的观点是一致的。
一句话总结
通过对前人研究的回顾,研究者发现:呼吸诱发震荡是一种全脑节律。啮齿动物一些脑区内,鼻腔呼吸诱发的神经震荡频率与呼吸频率相同。啮齿动物呼吸诱发节律与呼吸-γ耦合相关研究启示我们:同时记录呼吸和电生理数据对正确识别大脑振荡是必要的,呼吸诱发的振荡有助于大脑中的远程信息传递。这些结论也可推论到人类中。
原文
Tort, A. B. L. , BrankacK, J. , & Draguhn, A. . (2018).Respiration-entrained brain rhythms are global but often overlooked. Trends in Neurosciences

微信扫码或者长按选择识别关注思影
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
第十四届磁共振脑网络数据处理班(重庆,7.26-31)
第三十届磁共振脑影像基础班(南京,7.31-8.5)
第十届脑影像机器学习班(南京,6.30-7.5)
第十二届磁共振弥散张量成像数据处理班(南京,6.18-23)
第二十届脑电数据处理中级班(重庆,8.9-14)