

本文从设计和解释心理学实验的角度介绍了我们对TMS机制的理解现状。讨论了使用TMS进行行为研究的进展,总结了TMS在心理时间测量、感知、意识、学习和记忆方面的不同用途和挑战,是经颅磁在心理学上应用的经典好文。
1 前言
第一波人类经颅磁刺激(TMS)的行为研究,在试图产生类似神经心理学损伤效果和我们对这种方法的基本生理学的理解之间存在着脱节。由于运动诱发电位(MEP)的可靠性和可操作性,对运动皮质的研究引领了生理学理解的进程。当心理学家试图模仿神经心理病人的症状,或者测试有关过程的时间或顺序的理论时,我们必须尝试将对TMS在初级运动皮层(M1)上的兴奋和抑制作用的理解与自己的研究相结合,这并不容易。如果我们复制出一种临床症状,比如语言缺陷、空间忽视、注意力受损,面容失认症,或计算障碍,是否能够证明受刺激区域是行为的关键区域,是否是由于电流传播而无意中刺激了邻近区域,或者是否是二次刺激了另一个解剖上相连的远端区域?根据刺激部位的实验对照、时间测定和任务的特殊性,有充分的依据来推断解剖和生理效应,许多发现很有趣,而且在神经心理学意义上具有表面效度,但要从神经意义上解释这些效果是非常困难的。
问题是,M1以外的大多数脑区没有提供类似于MEP的可衡量的指标。有人把第一波TMS认知研究称为“点击式时代(point-and-shoot era)”,即确定一个假定的神经心理学感兴趣区,对其进行刺激,目的是扰乱与其相关的功能。我们发现了许多有趣的和可复制的效应,尽管时间有限,但它们仍然存在且被多次研究,但缺乏可信的机制解释限制了这些发现的影响。
为了使行为TMS的发现具有生理学证据,我们提出了三种机制。第一个是TMS对认知功能的影响是由神经噪音引起的,TMS导致了刺激区无组织的活动,阻止其正常功能的发挥。第二是相互竞争,TMS抑制了神经活动,即通过减少信号而不是增加噪音。第三,Schwarzkopf认为行为增强和损伤是因为TMS诱导了随机共振效应。然而,这些建议仍然是未经检验的假设。
2 TMS与心理学新时代的五大因素
该领域的成熟基于五大支柱的进步:
第一个是TMS引起的生理机制和行为变化之间的联系。我们重点介绍了TMS和非人灵长类动物的单细胞记录的工作;
第二个是使用TMS和脑成像来理解人类认知网络;
第三个是从运动系统中的峰时依赖可塑性(STDP)研究中采用和发展配对关联刺激(PAS),以及认知研究中双线圈皮质-皮质PAS(ccPAS)。
第四个是状态依赖,与TMS实验制定、执行和解释息息相关;
最后一个是行为,使用TMS来提高对心理功能的理解,并测试认知过程理论。
3 生理和行为
在介绍TMS时,最常收到的问题是,您如何知道它在做什么?它的传播距离有多远?它还会中断网络的其他哪些部分?在神经元水平上发生了什么?效果会持续多久?如果我们考虑TMS的单个脉冲,我们现在就可以对这些问题做出良好的回应。
传播始终是一个推论性问题,其依据是,如果线圈移动几毫米或旋转到不同的角度,MEPs或行为效应将会减弱或消失。模拟TMS的传播过程是困难的,因为脑脊液和白质/灰质具有不同的传导性,当然模型也不涉及行为状态。
阐明这些问题所需的方法是直接的生理记录。最初,这些记录是在麻醉状态中进行的。从这些研究中,我们了解到TMS在两个阶段具有特定的兴奋或抑制效应(即抑制之后是更长的兴奋时间);成对脉冲TMS在M1中的效应反映在视觉皮层的诱发电位活动中,并且TMS诱发电位活动是状态依赖的(与刺激前的神经元活动水平有关)。但要想在生理和行为之间建立联系,需要对清醒的、行为正常的猴子进行记录。Ortuno刺激执行视觉检测任务的猴子的初级视觉皮层(V1),并记录外侧膝状核(LGN)的活动,TMS诱导的LGN活动变化证明了次要位置的活动受到影响,其影响是状态依赖的。
最全和最新的进展来自非人灵长类动物的单细胞记录,在视觉线索提示下的伸展、抓取、举起和保持任务训练中记录数据。在一些试验中,猴子的PFG上接受了运动阈值(MT)60%或120%的单脉冲TMS。记录取自在任务中使用手的对侧的下顶叶PFG的500多个细胞活动,作为TMS强度、刺激线圈的距离和任务相关性的函数进行分析(图1)。在MT为120%时,TMS对单个神经元有明显影响。TMS脉冲会产生伪迹,所以实验分析了脉冲后10ms的活动。TMS的主要作用是在前50ms诱发一连串动作电位。在与任务无关的神经元中,这与传递TMS的任务阶段(即提示、举起、抓取、保持)无关。在其他神经元,早期兴奋之后是一段抑制期,在少数神经元中,第三阶段的兴奋可以持续长达250ms(平均小于100ms)。对线圈距离的分析和与相邻神经元的比较表明,TMS的影响仅限于2-3 mm。在MT的60%时,TMS几乎没有生理效应。
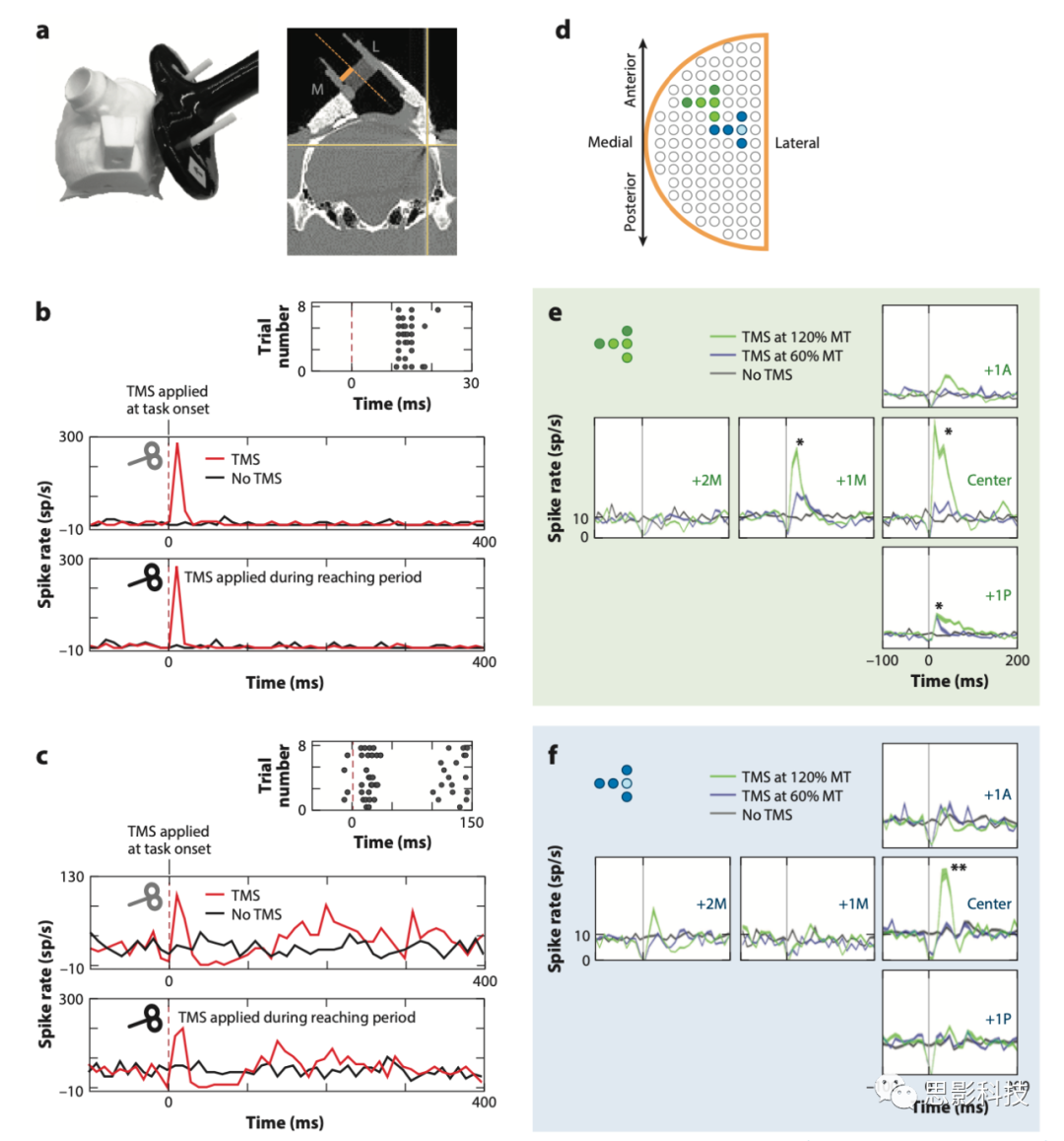
图1 TMS对猕猴顶叶皮质区单个神经元的影响
注释:(a)猴头部和TMS线圈的3D模型,显示记录电极的接近角度。
(b)在伸手抓握任务期间,当运动阈值(MT)为120%时,单细胞对TMS的反应。无论是在任务开始时(灰色符号)还是在到达期(黑色符号)施加TMS,刺激均在约10至40 ms之间诱发一波活动(红色曲线)。在非TMS试验中,活动无变化(黑色曲线)。光栅图举例说明了响应的一致性。
(c)另一类反应是抑制-兴奋-抑制。该神经元的反应显示与b组中的细胞相同的初始兴奋性爆发。当在任务开始时应用TMS(灰色符号和插图光栅图)时,有三个活动阶段:兴奋(0–40 ms)、抑制(40–100 ms)和兴奋(100 ms+)。在某些细胞中,较长的活性维持期可延长至300 ms。黑色曲线显示了非TMS试验中神经元的活性。(d)两只猴的记录位点网格,显示记录的空间特异性。
(e,f)该图显示了在TMS线圈中心(中心)、1 mm前部/后部(+1A,+1P)和1或2 mm中部以下的五个记录位点的平均细胞应答率(+1M,+2M)。这些图表显示了在120% MT下对TMS刺激的反应(绿色曲线)、在60% MT下对TMS的反应(紫色曲线)以及没有TMS刺激时的反应(灰色曲线)。
这项实验的最后一个发现是单脉冲TMS也在刺激区的中心和外围诱导了低频振荡的增加。这些振荡是TMS的诱发电位和长时程增强效应的因素,大多在δ和较低的θ范围。在脑电研究中,这种振荡没那么显著。然而,单细胞反应和脑电的空间和时间尺度不同。单细胞水平上的相对局部低频振荡与脑电、脑磁图(MEG)中频率变化较大的全局频率之间的关系是今后研究的一个方向。
从心理学的角度来看,也许最有趣的发现是,在一些神经元中,第一阶段记录的瞬时脉冲和第二阶段活动的减少与抓取任务表现受损的情况一致。已知PFG中的神经元参与行动目标选择和抓取行为。这是迄今为止将TMS诱导的神经活动变化与对行为的影响直接联系起来的最强有力的证据。
Romero也开始评估持续θ脉冲刺激(cTBS)对单细胞反应的影响。他们测量了300次cTBS脉冲后的反应,发现PFG区的大多数细胞活动明显减少。反应分为三组:早期组,47%的细胞在cTBS后的前5分钟受到影响;后一组,44%的细胞在cTBS后5-30分钟表现出抑制效应;以及非常晚的一组剩余细胞,在cTBS后30分钟以上兴奋性降低。这些结果表明,θ脉冲刺激虽然基于动物研究并广泛应用于行为研究,但并不具有单一的效应,这一点在解释行为研究时必须加以考虑。
这些发现是我们理解TMS生理效应的一个里程碑,至少为回答有关局部扩散、与任务相关的细胞相互作用以及效应持续时间提供了起点,还有助于解释神经噪声、信号减少和随机共振等机制。早期和晚期的兴奋性反应(图1)对应于噪音效应,减少响应的第二阶段正好是信号减少假说所预测的。振荡阶段,与早期脉冲和抑制阶段不同,发生在阈下刺激,这与随机共振描述是一致的。
4 功能解剖网络
TMS与功能磁共振成像的结合使我们能够在整个大脑中描绘出神经损伤的远程影响。这种方法的价值有三个方面。首先,它使我们能够描绘TMS的次要影响。其次,任何复杂的人类行为都是通过分布式脑网络进行计算的,将TMS与功能磁共振成像相结合,可以绘制出这些网络的范围和功能,以及因果关系。第三,存在无法直接用TMS刺激的皮质和皮质下区域;但是,TMS-fMRI联合研究表明,刺激侧面脑区可以调节偏远区域的神经反应,包括海马旁回和杏仁核。
研究额叶是如何自上而下地控制视觉感知和决策的,这是一个经证实非常有用的TMS-fMRI研究。在视觉辨别任务中额叶参与自上而下控制,在对额叶眼动区(FEF)进行TMS刺激的同时记录fMRI数据,发现刺激右侧FEF,V1至V4区域中央视野的视网膜视觉拓扑表征的活动降低,外围视野的表征增加。一项后续的TMS研究证实了这些结果的行为学相关性,证明了TMS刺激FEF,增强了外围视觉刺激相对于中心视觉刺激的感知对比度。
TMS-fMRI联合研究在研究其他认知网络方面也是非常有价值的。之前令人困扰的问题,即行为影响是否源于二次刺激,变得容易解决。人脸感知网络就是一个很好的例子。研究表明,不同的脑区优先代表不同的面部特征,如情绪、身份、注意点和社交线索。TMS-fMRI可以系统地干扰这些脑区,并检查网络连接的因果关系。Pitcher使用这种方法来区分处理动态和静态面部特征的皮层通路。通过θ脉冲 (TBS) 刺激右侧枕叶人脸处理区(rOFA)或右侧颞后上沟(rpSTS)。与现有的人脸处理模型相反,研究发现,静态面部特征(如身份)和动态面部特征(如表情)是通过从早期视觉皮层开始的分离的皮层通路进行处理的。
像OFA和pSTS这样的脑区可以被直接刺激,人脸感知网络的其他部分,如杏仁核,不能直接成为TMS的目标,但可以通过二次刺激的网络来探究。非人灵长类的神经解剖学研究确定了一条从STS到杏仁核的皮质通路。Pitcher将离线TBS和功能磁共振成像结合起来,被试在观看面部和物体视频时进行扫描,通过对rpSTS进行TBS刺激,右侧杏仁核对面部视频的神经反应减少,这一结果与非人灵长类数据一致(图2)。
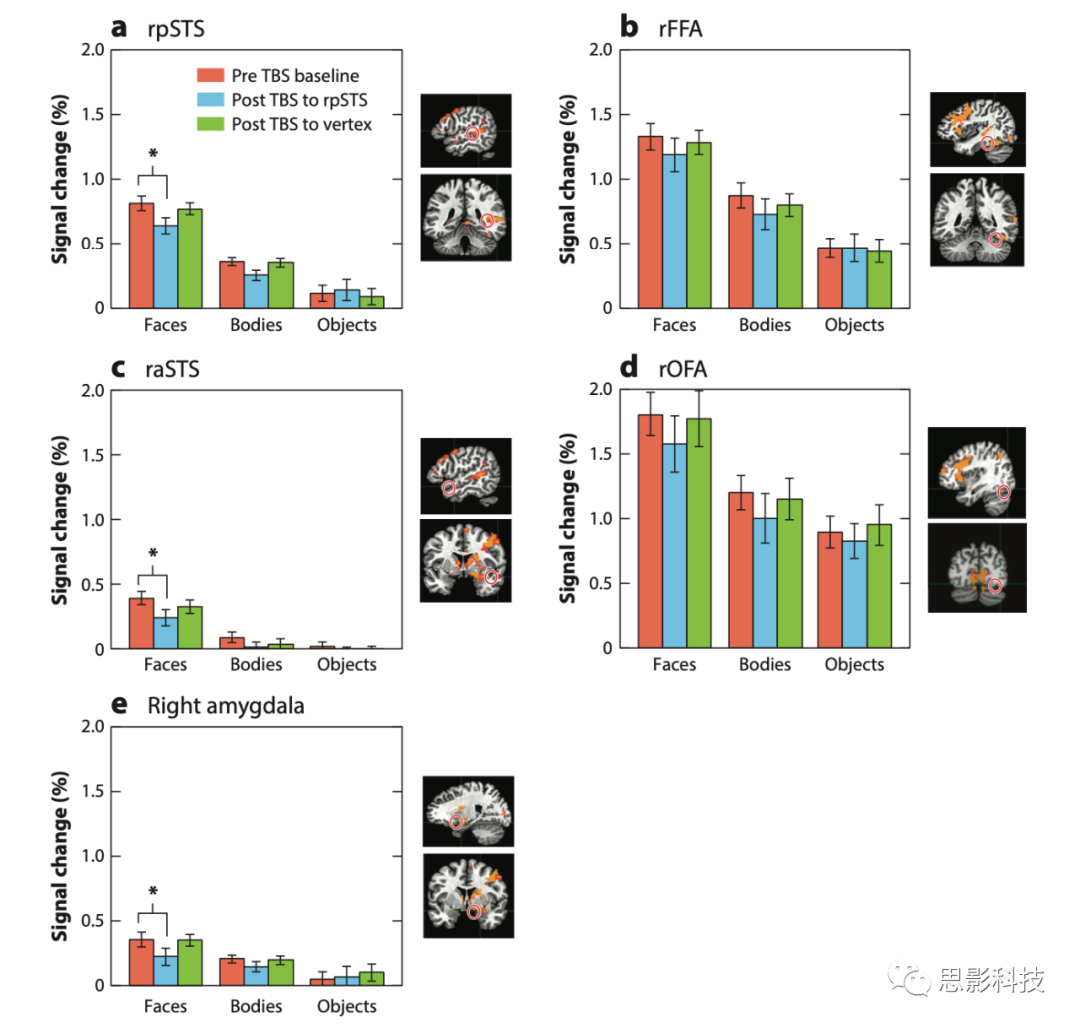
图2五5个ROI在cTBS前后动态面部、身体和物体刺激的信号变化
进一步可以使用TMS-fMRI在没有任务要求的情况下观察认知网络。Handwerker结合TMS和静息态功能磁共振成像(rsfMRI),绘制了整个人脸感知网络被短暂破坏的远程影响。与控制组相比,在rpSTS施加TBS刺激,静息态网络连接减少,不仅在rpSTS和其他面部选择区之间观察到,而且在大脑腹侧、内侧和外侧表面观察到(如,右侧杏仁核和双侧梭状回和枕叶人脸处理区)。在没有任务需求的情况下观察网络完整性,使TMS-rsfMRI具有临床价值,因为被试在扫描仪中完成任务很困难。例如,TBS结合rsfMRI已被用于研究衰老,视觉空间忽视、小脑连接和抑郁症。
结合rsfMRI和TMS也可以用来测量刺激如何改善解剖网络中的功能连通性。Wang研究了参与联想记忆的皮层-海马大脑网络,使用rsfMRI确定左侧海马和左侧顶叶之间的功能连接。连续5天在左侧顶叶部位持续高频TMS(20 Hz)刺激20分钟。与假刺激相比,TMS增加了刺激部位和左侧海马之间的功能连接,伴随着联想记忆任务中行为表现的改善。
5 峰时依赖可塑性
TMS效应的基本生理学和TMS与fMRI相结合所揭示的网络反应,证实了心理学实验中关于TMS的重要推论。使用两个线圈刺激大脑皮层的研究,在生理和行为学上也取得了很大进展,即STDP(峰时依赖可塑性),指的是突触前和突触后活动随时间的变化。这一Hebbian机制已经在小动物上进行了研究,神经药理学研究揭示了N-甲基-D-天冬氨酸(NMDA)和谷氨酸在诱导长时程增强中的作用以及GABA突触在抑制中的作用,具有显著的时间特异性。Hebbian学习模型是,重复成对刺激一个脑区(条件脉冲),几毫秒后再刺激第二个脑区(测试脉冲),提高了两个部位之间的突触效率,使得第二个脑区更容易接受来自第一个脑区的输入。在人体中,使用TMS对运动系统的研究表明,使用外周神经刺激与皮质TMS脉冲配对的PAS,如果这两个刺激相差几毫秒的时间窗,可以诱导MEPs变化。刺激正中神经(MN)10ms后,在M1施加TMS刺激降低了随后的MEP,而25ms的MN-M1间隔增加了MEP。Rizzo称之为ccPAS(皮质-皮质PAS),我们使用这个术语来指代所有关于可塑性的双皮质刺激研究。在右手(左半球) 和左手(右半球)的M1上以0.05 Hz施加TMS脉冲30min,两个半球之间的延迟为8毫秒。为每个被试设置刺激强度,范围在静息运动阈值的115%到125%之间。生理效应是增加右半球手部表征的MEP,降低大脑半球间抑制(IHI)。通过在左右M1上施加35-45毫秒的长延迟的单个脉冲来测量IHI,这些脉冲会抑制MEP。在行为学上,ccPAS加快了左食指(右半球)的反应时间长达30分钟。
ccPAS(皮质-皮质配对关联刺激)的M1效应似乎是稳健的和可复制的。Buch在被试完成伸展和抓握任务时,在腹侧运动前区(PMv)施加TMS以调节M1。持续至少一个小时的STDP(峰时依赖可塑性)效应是状态依赖的,因为当被试执行任务时,PMv上施加TMS诱导了M1的兴奋,但静息态时,PMv上施加TMS抑制了M1的活动。然而,这对任务本身的表现没有影响。Chao在后顶叶皮质(PPC)上施加TMS后8毫秒调节M1,获得了M1兴奋性的变化。他们发现STDP对MEP的影响持续了一个多小时,但对任务没有影响。Johnen使用ccPAS探究了在PMv和M1之间诱导更强的连接,是否会转化为在被试执行伸展抓取任务时通过fMRI测量的功能连接的增加,发现ccPAS在任务执行期间增加了两个脑区之间的连接强度,而没有增加静息态的连接。随后的分析表明,PMv-M1的ccPAS对感觉运动网络中其他节点之间的相互作用具有特定的影响。PMv与AIP之间的功能连接增强,而背侧运动前区(PMd)与M1、PMd与后顶上小叶(pSPL)之间的功能连接降低。对行为学没有影响。
Santarnecchi还研究了ccPAS(皮质-皮质配对关联刺激)如何改变脑网络活动(图3)。他们将ccPAS应用于默认网络(DMN)的顶叶和任务积极网络(TPN)的前额叶。根据fMRI研究,这两个脑区在各自网络中的连接强度是负值。根据先前对额顶网络的STDP(峰时依赖可塑性)研究,采用三种时间条件:DMN/TPN ISI +10ms,DMN/TPN ISI -10ms,DMN/TPN ISI=0ms。在15分钟内以0.2 Hz的频率施加成对的脉冲。他们没有测量运动皮层中的STDP,而是研究当因变量为BOLD信号时,认知网络的不同部分对TMS脉冲有何反应。两个发现是:(a)当DMN脉冲先于TPN脉冲10ms时,两个脑区的负连接性发生了明显的变化,以及(b)在三分之一的被试中,连接变为正值(图3)。其影响也是依赖于状态,也没有任何行为学影响的报道。

图3 ccPAS对左角回和左额叶之间的功能连接强度的影响
我们对ccPAS(皮质-皮质配对关联刺激)感兴趣,是希望STDP(峰时依赖可塑性)在临床上应用,在Rizzo之后的研究,尽管STDP发生了显著的生理变化,但大多数研究要么没有测量,要么没有看到STDP带来的行为变化。Kohl尝试将ccPAS诱导的STDP的生理与行为相匹配。他双向刺激了下额叶(IFC)和前SMA。他们假设在IFC之前4或10ms刺激前SMA,会抑制丘脑底核(STN)并干扰行为任务,而在刺激前SMA之前 4或10ms刺激IFC会促进STN和任务执行。他们以0.2 Hz的频率施加了100对ccPAS脉冲。他们给被试的两个任务是停止信号反应时间(SSRT)任务和时间折扣任务。虽然他们报告了对SSRT的一些影响,但没有发现对延迟折扣任务的影响。这一领域的挑战是建立生理上STDP效应(通过MEPs或fMRI测量)和行为之间的联系。
Veniero诱导了长时程增强(LTP)和长时程抑制(LTD)的STDP(峰时依赖可塑性),并测定其对诱发电位的影响。通过以5毫秒的ISI在8分钟内用100个脉冲对刺激PPC和M1来诱导双向STDP。当ccPAS对MEP有抑制作用时,PPC和M1之间的β相干性增强。当ccPAS诱发MEP的兴奋时,两个区域之间的α相干性增加。Veniero使用的范式,与结合TMS-EEG在视觉、注意和意识方面的工作是一致的,值得推广。
另一个问题是产生行为效应。Momi使用ccPAS同步刺激左额顶网络的两个节点。用2 Hz、ISI为10ms的脉冲对双向施加ccPAS,时间为15分钟。经过5周的重复刺激和行为测试,观察到STDP和方向性行为改变的迹象。额-顶叶ccPAS与关系推理的改善有关,而顶-额叶ccPAS与逻辑推理任务的改善有关。这是一个罕见的ccPAS(皮质-皮质配对关联刺激)产生复杂行为变化的例子。
6 状态依赖关系
从某种意义上说,所有的TMS研究都是基于状态依赖的。感知和认知实验中的状态依赖方法主要归功于Juha Silvanto的努力。TMS的作用取决于被刺激的脑组织的当前兴奋状态。然而,在认知实验中,背外侧前额叶皮质(DLPC)、PPC、FEF、OFA等的当前状态,没有诸如MEP的直接测量指标。Silvanto的主要贡献是利用心理实验中的状态依赖来提高TMS的功能分辨率。
Silvanto利用适应性来影响被刺激区域的初始状态。他的第一项研究可以作为认知依赖状态的TMS模型。被试适应颜色/方位刺激30秒,并报告测试刺激的颜色。在呈现测试刺激后的不同时间给予TMS。在非TMS试验中,被试观看适应刺激的补色,但在视觉区施加TMS诱导了对原始的适应刺激颜色的偏向。
该方法现已广泛应用于心理意象的研究,短期记忆、字母选择、抽象、动作观察和语义处理。Cattaneo的研究观察到了在视觉或语言区之外施加TMS产生的命名效应。他们测试了TMS对左侧PMv,工具词编码中类别特异性神经元表征的影响。给被试呈现一个初始类别名称(“工具”或“动物”),以使PMv适应一个或另一个类别的对象,然后当目标词与初始类别一致或不一致时施加TMS,初始效应消失了。状态依赖的TMS的另一个例子来自对动作识别的研究。Mazzoni使被试适应情感运动的点光显示。他们在点光源显示开始时,在前顶内沟(aIPS)、pSTS或枕极上施加重复TMS(rTMS)。只有在aIPS上施加TMS,才能影响被试对快乐或恐惧运动的识别。
我们可以把STDP和依赖于状态的TMS这两个最新进展结合起来。有三项研究可以作为状态依赖和ccPAS如何结合使用的例子。第一个使用三个线圈ccPAS刺激双边M1和SMA。它们诱导了双向和时空特异性STDP,当SMA-M1间隔6ms时MEP增强,而M1-SMA间隔15ms时MEP降低。为了使这些联系发生,左M1和右M1必须同时受到刺激。第二项研究,Romei在V5和V1施加TMS,以0.1 Hz的频率传送90对ISI为20ms的刺激,改善了被试在运动一致性任务中的表现。后续研究中,Chiappini表明这种强化效应取决于状态(图4)。
一类特殊的实验使用双线圈刺激来探测脑区之间的相互作用。Hartwigsen实验室的研究人员称它们为条件干扰实验,并展示了它们在动作编程和语义决策方面的作用。Hartwigsen在被试执行空间预先提示的反应时间任务之前,使用两个线圈在PMd上施加1 Hz离线TMS。任务中,在颞上回(SMG)上施加10 Hz在线rTMS。这两个脑区的联合刺激使无效线索试验时反应时间减慢,但单独刺激SMG不会产生减慢。他们将其扩展到不同形式的离线刺激和不同的行为领域。在被试执行语音或语义决定任务时,在SMG或角回上施加cTBS作为离线刺激,在线条件下在前额叶或后侧下额叶(IFG)上施加10 Hz刺激。离线角回和在线aIFG刺激使语义判断任务的RT减慢。
7 行为发现
视觉意识一直是许多TMS研究的主题。Allen使用离线cTBS再现了失明效果。θ刺激可能不会在单个部位产生单一效应。通过将cTBS与磁共振波谱(MRS)和MEG相结合,Allen的研究表明,尽管cTBS降低了皮层活性,增加了GABA浓度,但意识阈值降低了,而不是像预测的那样提高了。效应在cTBS后0到48分钟内是稳定的。他们提供了一个很好的例子,说明了生理抑制性TMS是如何导致行为增强的。同一研究小组还使用脑磁图作为使用TMS的计时指南,以测试前SMA和IFC在反应抑制中的作用。
离线刺激后行为改善的一个例子表明,使用心理物理任务可以帮助解释生理机制。V5上施加TMS通常被用来干扰运动知觉,但由于其较大的感受野,该区域也涉及到空间抑制。Tadin在V5或后枕叶上施加1 Hz的TMS 15min,同时被试执行运动辨别任务,造成的抑制中断导致了运动辨别任务的改善。
顶叶的注意功能长期以来一直是TMS研究的焦点。Rangelov探究了前馈视皮层输入对注意力捕获是否必要。他们绕过了视静脉输入,将TMS作为有效和无效的信号来诱发光幻视。与真实的视觉刺激相比,产生了相似的效果,尽管略弱,但绕过解剖通路的范式可以推广至TMS-fMRI、TMS-EEG和状态依赖实验。Herring的研究表明,单脉冲TMS诱导的视觉系统振荡活动与内源性振荡具有相同的神经起源和特征。他们测量了被试在跨模态注意任务中的经颅诱发电位(TEP)和脑电图α反应。在视觉注意条件下,TMS增强了TEP(N40)的一个早期成分,而α功率降低。
顶叶除了注意力还涉及其他功能。Vesia基于非人灵长类动物的单细胞研究解析了顶叶的功能特异性(图5)。他们在上顶叶枕区(SPOC)和顶内沟的两个位置(顶内沟和角回)施加了三个10 Hz的TMS脉冲。被试执行延迟的空间扫视或到达任务,rTMS在延迟期内施加。顶内沟和角回上的TMS增加了到达反应的变异性,对对侧目标的到达和扫视准确性降低。在SPOC上施加TMS使到达任务的准确性降低,但不影响扫视。
在记忆领域,TMS的研究范围从启动记忆到工作记忆、情景记忆、元记忆和陈述性记忆。传统意义上,记忆与海马和额叶有关,但TMS的研究也发现了枕叶的记忆功能。Hilbert研究数字广度的工作记忆时发现,在任务开始前,对枕极施加近10分钟的1Hz的TMS会损害被试的记忆表现。几项TMS研究发现感觉皮层在工作记忆中的作用,例如在V5上施加在线rTMS。Van Lamsweerde用单脉冲TMS在枕叶上可以扰乱被试能记住的项目数,可能是干扰了工作记忆的巩固。他们还肯定了在研究记忆的某些方面时考虑感觉皮层刺激表征的重要性。
在情景记忆领域,一系列研究使用rTMS、rsfMRI、fMRI和EEG来区分不同的记忆路径,并在24小时内增强情景记忆。第一个实验在面孔-单词联想记忆任务中改善行为。一项结合TMS和EEG的随访研究复制了这一发现,使用了物体位置记忆任务。TMS提高了空间精确度,但不是整体精确度,还引起了电生理测量的变化。成功回忆试验的θ-α范围的功率降低,TMS刺激后回忆相关事件的事件相关活动电位的幅度也降低了。还有研究使用功能磁共振成像测量了TMS对记忆形成的影响。通过三项功能指标和三项不同的记忆任务发现,在后侧皮质-海马网络施加rTMS选择性地改善了记忆,并改变了整个网络的活动模式。
8 TMS对行为影响的机制是通用的?
在状态依赖实验中,我们看到TMS激活了适应性的神经元群体,优先激活当前任务相关的神经元群体。因此,在物理参数空间、时间、相关性和意识状态之间,有一个共同的特点,即TMS优先刺激弱激活的群体。我们首先强调了更好地理解TMS生理学的必要性,但这些行为实验为未来的生理记录提出了新的问题。如果像过去一样,我们被问到如何知道TMS在做什么,我们可以把这个问题返回给有心理背景的电生理学家,然后问,心理状态对生理反应有什么影响?了解TMS的生理效应和被刺激的大脑的生理状态之间的双向相互作用是下一阶段进展的先决条件。
9 总结
本文作者认为,我们对过去10-12年的进步可能过于乐观,但诸如单细胞记录、STDP、状态依赖、解剖可靠性、MEP、光幻视和网络效应等研究仍是有参考价值的。现有研究已经在技术上、概念上、和经验上取得了进展。但是,与纯粹的行为实验相比,将TMS与各种方法(fMRI、EEG、MEG、MRS、眼动、MEP等)相结合,样本量往往很小。而且使用两个线圈并不容易,且没有报告效应量大小,很少有预先注册的研究。所有这些都是弱点且是可以改进的:报告无效结果,考虑实验的预注册,充分报告刺激方法,采用和改编来自其他研究领域的开放数据实验。我们鼓励所有作者和审稿人参与提高该领域的整体研究质量。
如需原文请添加思影科技微信:siyingyxf或者18983979082获取,如对思影课程及服务感兴趣也可加此微信号咨询。觉得对您的研究有帮助,请给个转发,以及右下角点击一下在看,是对思影科技莫大的支持。

微信扫码或者长按选择识别关注思影
非常感谢转发支持与推荐
欢迎浏览思影的数据处理业务及课程介绍。(请直接点击下文文字即可浏览思影科技所有的课程,欢迎添加微信号siyingyxf或18983979082进行咨询,所有课程均开放报名,报名后我们会第一时间联系,并保留已报名学员名额):
第三十六届磁共振脑影像基础班(南京,2021.1.6-11)
脑磁图(MEG)数据处理学习班(预报名)
数据处理业务介绍:
思影科技拜访式培训服务
思影科技EEG/ERP数据处理业务
思影科技脑电机器学习数据处理业务
招聘及产品:
招聘:脑影像数据处理工程师(重庆&南京)