

染色体22q11缺失是最常见的微小缺失综合征。发病率为1/2000至1/4000。它可引起一系列的临床障碍。最新的研究表明,患者有精神行为异常者高达60%。从目前已有的研究来看,该疾病的表型异质性高,引起的临床症状极为复杂。
在以往的单中心研究中,关于22q11DS的dMRI研究已经报道其存在白质微结构改变。但由于小样本和方法的不同致使结果存在异质性。以往的研究结果虽然都指出了22q11DS患者的白质微结构存在异常,但在具体部位和损伤程度上都难以达成结果的一致。因此,亟需大样本、多中心研究来提高该研究的跨中心可重复性以及结果的可信性。针对该问题,来自南加州大学凯克医学院的研究者使用了迄今为止最大的22q11DS研究样本进行了规范性的白质微结构研究,研究结果发表在Molecular Psychiatry杂志上。
该研究中,共包括了334名22q11.2缺失携带者和260名年龄和性别匹配的健康对照者;年龄范围6-52岁,这是迄今为止,在22q11DS患者研究中,由dMRI对白质微结构测量的一项最大的研究。
使用由ENIGMA-DTI工作组开发的处理方案,发现22q11DS中的平均、轴向和径向扩散率存在广泛降低,皮层-皮质和皮质-丘脑纤维最明显:放射冠、胼胝体、上纵束、丘脑后辐射以及矢状层(Cohen's d:0.9-1.3)。只有内囊后肢,其轴向扩散系数较高。22q11DS患者胼胝体和投射纤维平均FA比对照组高,但主要区域FA值比对照组低。22q11DS中的精神障碍与多脑区的扩散率显著降低有关。总体来讲,这些发现表明22q11.2缺失对白质微结构有很大的影响,尤其是在主要的皮层-皮质连接中。结合动物模型的发现,这种异常模式可能反映了外皮质层投射神经元神经发生的中断。
前言
22q11.2缺失综合征(22q11DS:也称为腭心面综合征或迪乔治综合征)由22号染色体长臂1.5-3巨碱基微缺失造成,是一种常见的染色体微缺失症状,在新生儿中的患病率为1/3000~4000。22q11DS与一系列的异常特征表现有关,如心脏缺陷、头面部异常及智能残疾等,尤其相较于普通人群增加了25倍的疾病患病风险。其中,与精神类疾病可能具有最特殊的关系,因为与发育迟缓相关的精神疾病风险增加了近乎3倍。值得注意的是,22q11.2缺失的小鼠模型中显示皮层2/3层的投射神经元神经祖细胞较少,导致皮层相关区之间的连接改变。因此,22q11DS是研究皮质环路发育障碍(如精神分裂症)潜在神经机制和遗传原因令人信服的模型。
dMRI能够非侵入性的测量白质微结构属性,FA作为DTI研究常用指标能够反应神经纤维的贯通性和密度。AD、RD指标在大量的脑部疾病中也发生改变。如低AD值反应轴突损伤和退化或轴突直径较小。在动物模型中,脱髓鞘化和髓鞘发育都可能导致异常RD值。MD值是细胞膜表面积与体积比的一种广义测量。
在22q11DS中经常报道白质微结构存在损伤,但迄今为止研究样本较小且研究结果不尽相同。例如,有些研究报道22q11DS相较于健康被试在有些大的白质纤维束存在FA值降低,如胼胝体、联络和投射纤维束,但也有少数表明FA值升高,或不同纤维束间高低不一。尽管部分22q11DS研究的结果混杂,但大多数报道一致发现了该类患者白质纤维的MD、AD和RD值的降低。因此,这种在宏观看起来一致,但在细节上存在较大异质性的结果阻碍了关于22q11DS中白质微结构异常的真实结论的形成。
已有研究已经表明,与精神疾病相关的白质差异在22q11DS中是有意义的,有研究发现,22q11DS中的精神症状与较高的FA值和低白质扩散有关,但不同研究间的结果脑区不同。此外,缺失断点存在变异性;85-90%的缺失个体有3Mb(A-D)缺失,包含46个蛋白编码基因,而10%的缺失个体有1.5MB(A-B)嵌套缺失。在22q11DS中白质差异差异部分可能是由于缺失大小不同,因为缺失大小影响皮质表面区域。
为了解决这些不确定性并确定影响22q11DS中白质异常的因素,ENIGMA-22q11DS工作组对10项独立研究的原始dMRI数据进行了一致分析并对组间差异和调节因子进行了Meta分析。解决如下问题:
(1)22q11.2缺失携带者和人口学匹配的健康被试之间的白质微结构是否存在一致的组间差异?
(2)两组间是否存在年龄效应差别,提示22q11DS中白质发育改变
(3)有精神障碍的22q11DS被试是否表现出更为严重的白质改变,并且这种差异与原发性精分是否具有重叠?
(4)缺失大小是否影响DTI指标?
(5)在22q11DS和健康被试中白质微结构改变与认知能力有关吗?
被试:
dMRI数据来自10家中心,作为ENIGMA-22Q11DS工作组的一部分进行分析。本研究纳入594名被试,334名22q11DS(年龄:16.8±6.43,女性:153名)260名健康对照(年龄:16.55±8.01,女性123名)人口学特征(如表1和表S2a,b)。
表1:一般人口学信息

表S2a:22q11DS信息
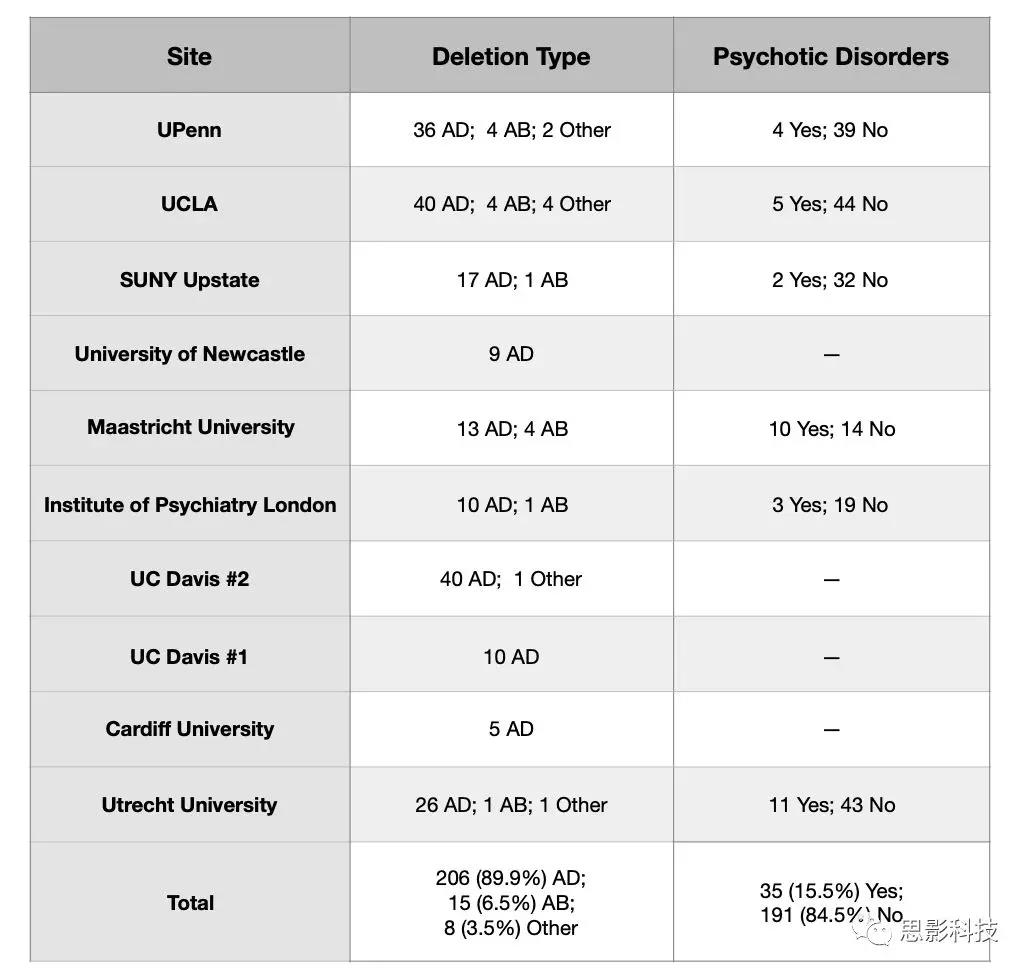
表S2b:伴或不伴精神障碍22q11DS信息

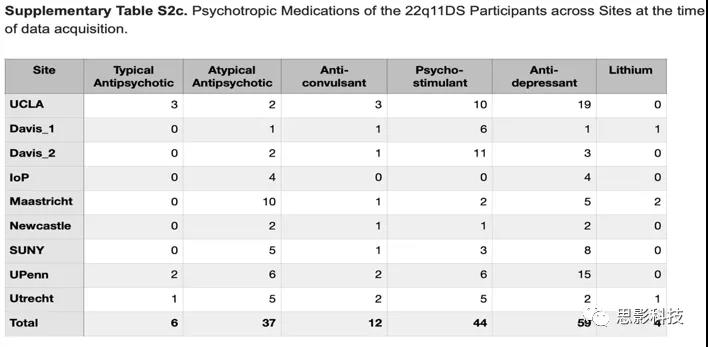
扫描时的精神用药情况如表S2c。个别研究详情见补充表S3(太长,查看原始补充材料)。所有研究参与者或法定监护人均已签定书面知情同意。
表S2c:各中心扫描时22q11DS的精神用药情况
![]()
样本详细基因型特性测量:所有中心在扫描时均进行了结构式的诊断访谈,确定终身精神病诊断。韦氏智商评估用于评估认知功能。用多重连接探针扩增技术(MLPA)测定缺失大小。本研究大样本允许独特地比较两种最常见的缺失类型(较长的A-D和较短的A-B,两种不同的基因缺失类型)对DTI结果的影响。从有MLPA数据的病例来看,206名受试者存在A-D缺失(89.9%),15名受试者(6.5%)存在A-B缺失(见补充表S3)。
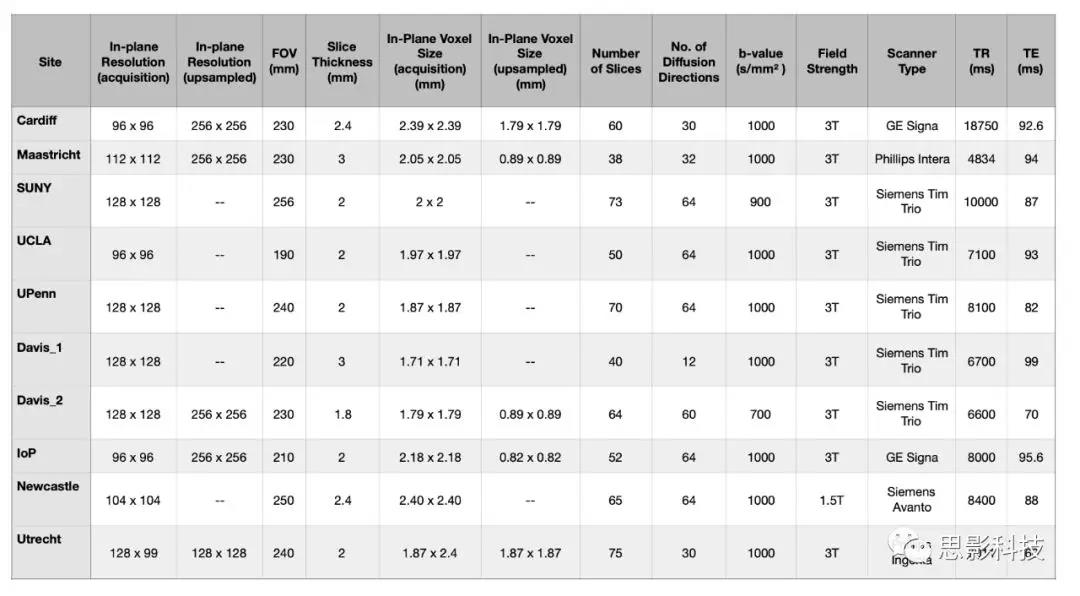
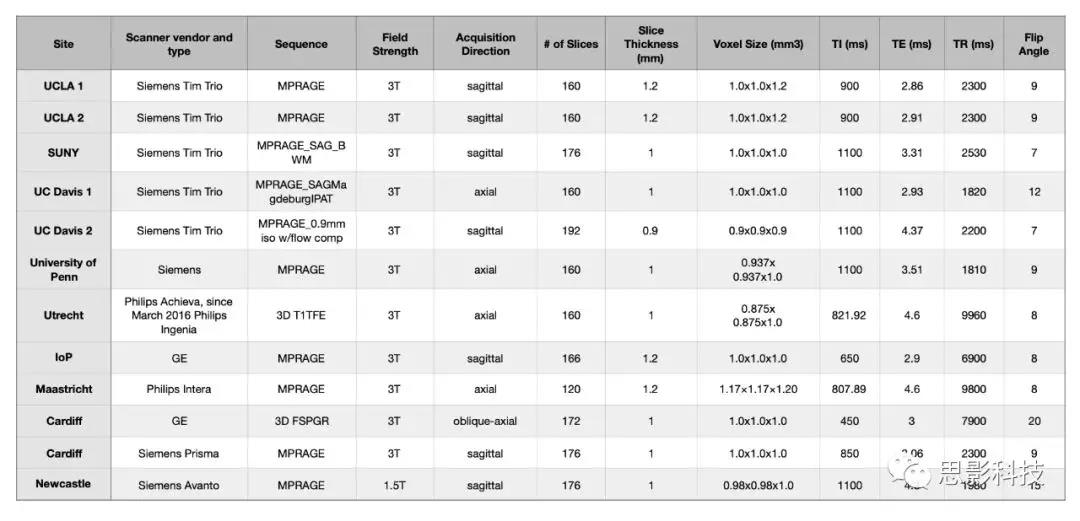
影像获取与处理:各中心dMRI和TI加权MRI扫描参数在补充材料(表S4和S5)中。表S4:dMRI参数
表S5:各中心T1参数
FA、MD、RD和AD地图按照ENIGMA-DTI流程中的描述进行骨架化,基于TBSS方法,确保所有数据都配准到ENMA-DTI模板。通过FSL计算基于JHU-ICBM-DTI-81模板的ROI骨架各参数平均值。在所有的分析中,每一指标使用双侧ROI左右值的平均值;包括了所有JHU-ICBMROI的平均值,排除了皮质脊髓束,中矢状穹窿区和扣带回束的海马部分,因为这些ROI很难进行可靠地配准,或在本研究队列中受到伪影影响。研究中包括的ROI如(图1)所示。
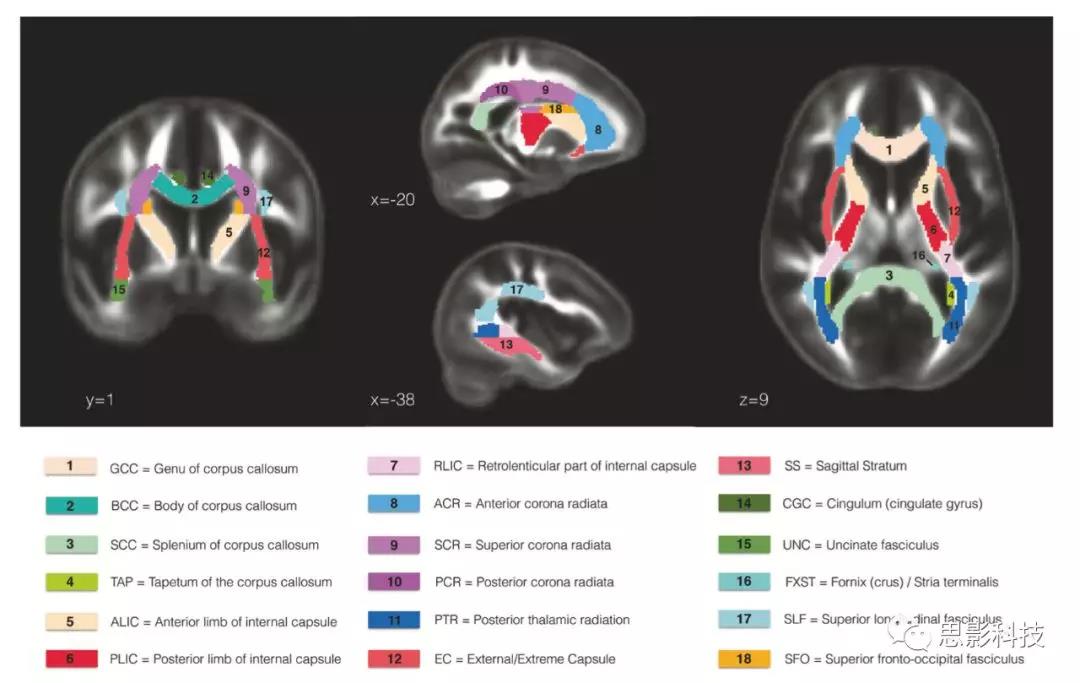
图1:JHU-ICBM白质图谱本研究ROI
22Q11DS和年龄对DTI结果的影响:
应用两种分析方法探究22q11DS和健康被试组间差异:Meta分析分别对每个中心进行统计比较,并将各中心汇总统计数据结合;Meta分析中数据统一且从个体被试汇集,对整个组进行统计分析。Meta分析包含来自8个中心9个独立数据集的540名被试:278名22q11DS(年龄:16.76±6.78)260健康被试(16.55±8.01)表1。因为Utrecht只包括22q11DS病例,所以未进入病例对照分析中。每个研究中心,进行线性回归分析,其中每个ROI的平均DTI指标为因变量,诊断为感兴趣的预测因子,年龄和性别作为协变量。鉴于获取的DTI指标,对于连合纤维和联络纤维的峰值在11到20岁之间,投射纤维的峰值在20岁早期时候,因此在模型中同时考虑了年龄的线性和二次效应。二次年龄项居中以避免与线性年龄项共线。此外,由于男女间在发育过程中DTI指标表现出不同的轨迹,因此模型中考虑了性别。计算诊断Cohen’s d效应大小。随后,进行逆方差加权混合效应Meta分析去结合个体位点效应大小。
因为多中心因素会影响DTI指标分布,因此,进行不同参数汇集dMRI数据研究时,对DTI指标进行额外的一致化是有利的。本研究应用了COMBAT算法对不同中心每个ROI的所有DTI指标(FA、MD、RD和AD)进行一致化处理。该算法使用经典贝叶斯框架来估计加性和乘性中心效应。以前曾用于多中心DTI数据的一致化,并且在建模和消除中心间变异性方面比其他几种方法更好。接下来,使用meta分析中测试的相同模型评估组间差异。最后,将年龄交互作用效应项纳入mega分析(分子进化遗传分析)模型,以检验22q11DS先证者相对于HC的年龄效应是否不同。
本研究使用Benjamini & Hochberg方法控制多重比较中的假阳性率。假阳性率阈值为5%(q<0.05)。计算每组模型的临界p值,具体:(1)meta分析;(2)mega分析;(3)mega分析,包括年龄交互作用诊断。
此外,鉴于先前健康个体(5-82岁)DTI指标非线性轨迹发现,本研究分别为每组(HC和22q11DS)每个白质ROI和每个DTI指标拟合了年龄的泊松非线性模型,以进一步研究年龄效应。使用先前一致化数据以减少中心的影响。正如Lebel等人研究,本研究测量了FA峰值年龄和MD、RD和AD最小值的年龄。两组均采用双尾t检验进行平均值剔除(α=0.05)。此后,计算了每个ROI从6岁(两组的最小年龄)到峰值/最小值,从峰值到46和52岁(分别为22q11DS和HC的最大年龄)的每一个DTI指标的百分比变化。采用带有bootstrap-t的Yuen方法比较了22q11DS组和HC组之间所有ROI的每一DTI指标百分比变化(α=0.05)。
精神障碍、染色体缺失大小及IQ对DTI结果的影响
为评估作为临床功能和遗传变异的白质结构的潜在差异。本研究在22q11DS被试中探测了精神障碍(精神障碍:35人,无精神障碍:191人)和染色体缺失(AD:206人,AB:15人)对DTI参数的影响。另外IQ作为组关联变量,本研究分别在两组被试中(22q11DS:304;HC:102)进行偏相关检验。在上述分析中ROI指标作为因变量,年龄、性别作为协变量,且进行FDR校正。
年龄和精神障碍起病时间具有强相关,并且在22q11DS组中患精神障碍和非精神障碍者间年龄具有显著差异(表S2b)。在22q11DS组中为评估精神障碍的效用,本研究应用局部非参ANCOVA检验方法。为检验22q11DS组精神障碍被试中观察到的白质微结构差异是否与原发性精分有重叠,本研究将22q11DS组中的结果与来自ENIGMA精分DTI工作组中的精分病例对照结果进行了比较,protocol与本研究一致。
跨中心的组别差异:
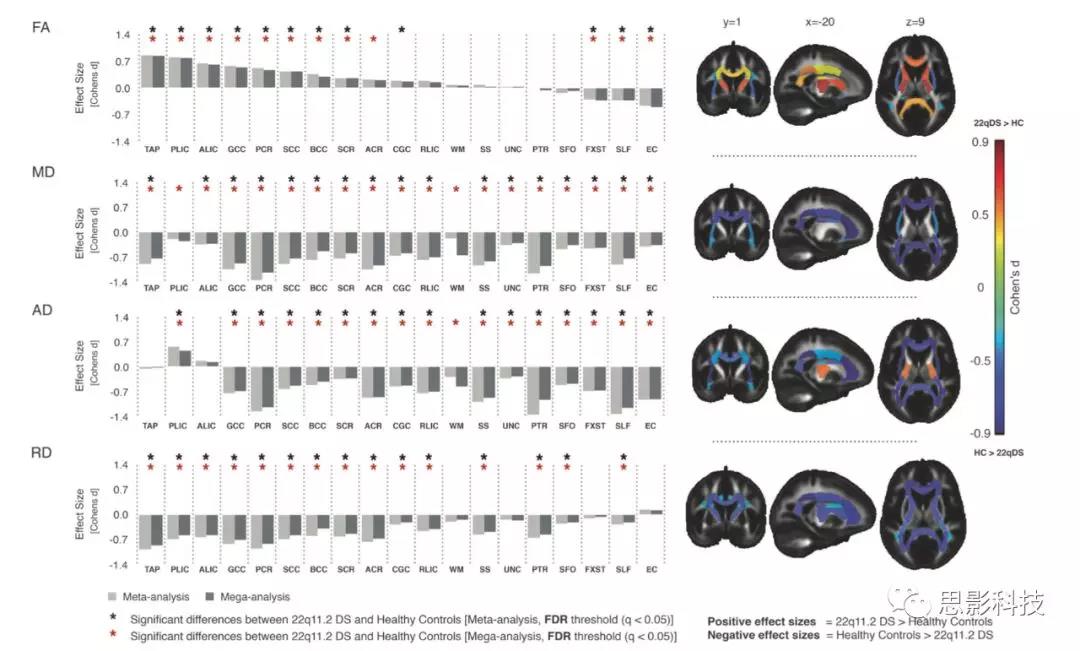
首先使用标准化处理流程探究22q11.2缺失携带者和HC之间白质微观结构是否存在一致的组间差异。同样重要的是确定数据的一致性是否允许对调节因素(精神障碍、缺失大小和IQ)进行进一步汇总分析。图2显示了从meta分析和mega分析得出的22q11DS与HC的组间差异:结果几乎相同,效应量相似。
![]()
图2:ENIGMA-22q11DS研究9个独立数据集的meta分析和mega分析
每个中心的效应大小如补充图1所示。在22q11DS和HC之间大多数ROI存在显著差异,在22q11DS受试者中扩散率值(MD、AD和RD)较低,但FA呈现混合模式。22q11DS患者在胼胝体毯(TAP)、膝(GCC)、体和压部(BCC/SCC)、内囊前和后肢(ALIC/PLIC)和放射冠后上方(PCR/SCR)的FA均显著高于HC,且效应中到大不等(d~0.3-0.8)。相反,与HC相比,上纵束(SLF)、穹窿/终纹(FXST)和外/外端帽状沟(EC)的ROI在22q11d显著低于HC(补充表S6和S7,过长,看原始补充材料)。
22q11DS受试者在几乎所有检测的ROI中MD值均显著低于HC组,在PCR和丘脑后辐射(PTR)中影响最大(d~1.0);两者都主要含有丘脑-皮质/丘脑-皮质和后脑区域皮质纤维。与MD一样,所有18个ROI,AD值在22q11DS中有15个较低。在22q11DS中只有PLIC处AD值表现出明显高于HC组。对于RD值,在22q11DS中,所有显示显著差异的ROI(18个ROI中的15个)均低于HC组,在胼胝体和PCR中效应最高(d~0.7)(补充表S6和S7,过长,看原始补充材料)。
如果您对DTI及其他模态脑影像数据处理感兴趣,请点击以下文字,或添加微信:siyingyxf进行咨询:
第六届任务态fMRI专题班(重庆)
第九届磁共振脑影像结构班(南京)
年龄相关效应:
鉴于年龄范围较大(6-52岁),本研究想确定22q11DS中的白质发育是否出现延迟或改变。如补充表S6所示,年龄对大多数ROI的所有指标都有非常显著的线性影响。FA与年龄呈正相关,而其他指标(MD、AD和RD)则为负相关。对于FA、MD和RD,几乎所有ROI中都存在显著的二次效应。在meta和mega分析中,AD显示的显著二次效应较少。然而,未观察到明显的年龄与诊断的交互作用(补充表S8,过长,看原始补充材料)。考虑到较大年龄的成人样本较少,本研究还对30岁以下的受试者进行分析,探索年龄与诊断的潜在作用,但结果与上述类似(补充表S9,过长,看原始补充材料)。
研究应用泊松回归模型,
以进一步评估年龄对白质发育的影响。模型并没有提供比上述线性回归模型更好的数据拟合。
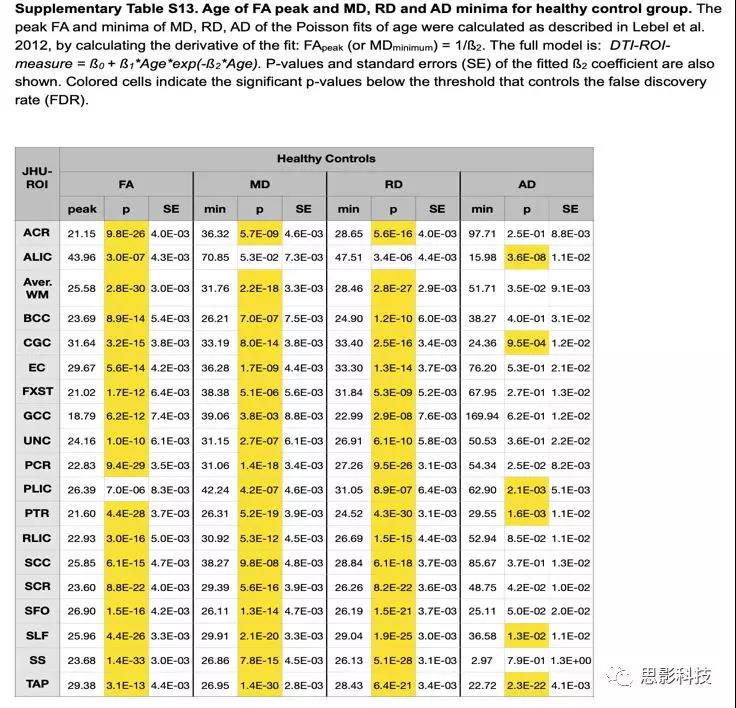
在所有DTI指标中,22q11DS组具有显著性峰值/最小估计值的ROI较少(见表S13和S14)。
表S13:HC组FA峰值,RD/MD/AD最小估计值年龄
表S14:22q11DS组FA峰值,RD/MD/AD最小估计值年龄
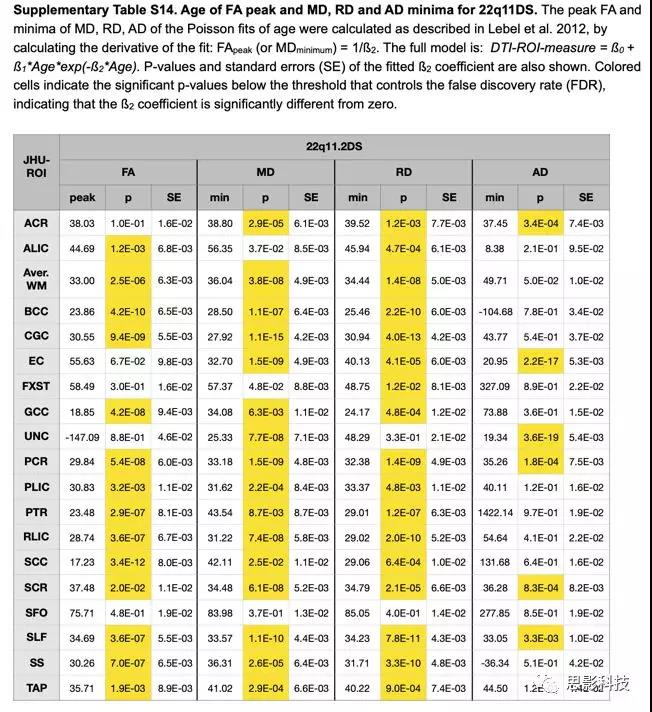
通常,那些没有显著性峰值/最小估计的ROI具有线性而不是指数增长和衰减轨迹。当比较HC和22q11DS之间的峰值FA(跨ROI)的平均年龄时, 22q11DS明显大。研究发现在22 q11DS中的最小RD时的平均年龄明显偏大,但最小MD和AD时的平均年龄没有差异。HC组在峰值后FA的平均百分比变化和最小RD和MD后的平均百分比变化也明显大于22 q11DS组,AD无差异(表S15)。
表S15:两组间FA峰值和弥散最小值差异

精神疾病的影响:
精神病患者中与染色体缺失相关的白质改变是否更严重?与没有精神病的22q11DS受试者相比,22q11DS受试中精神障碍者的扩散率总体较低,ALIC、PTR、扣带回(CGC)和SLF中的AD值明显较低。22q11DS精神障碍与GCC的RD和MD以及PLIC的MD显著降低有关。这些差异主要呈现在20岁至26岁之间的大多数ROI中;一些ROI(ALIC、PTR和SS)在16-17岁时表现出差异(图3和表S16:太长)。总体来讲,研究结果证实了由DTI指标反应的白质差异在22q11DS精神障碍患者中更为严重,在年轻人中更为明显。
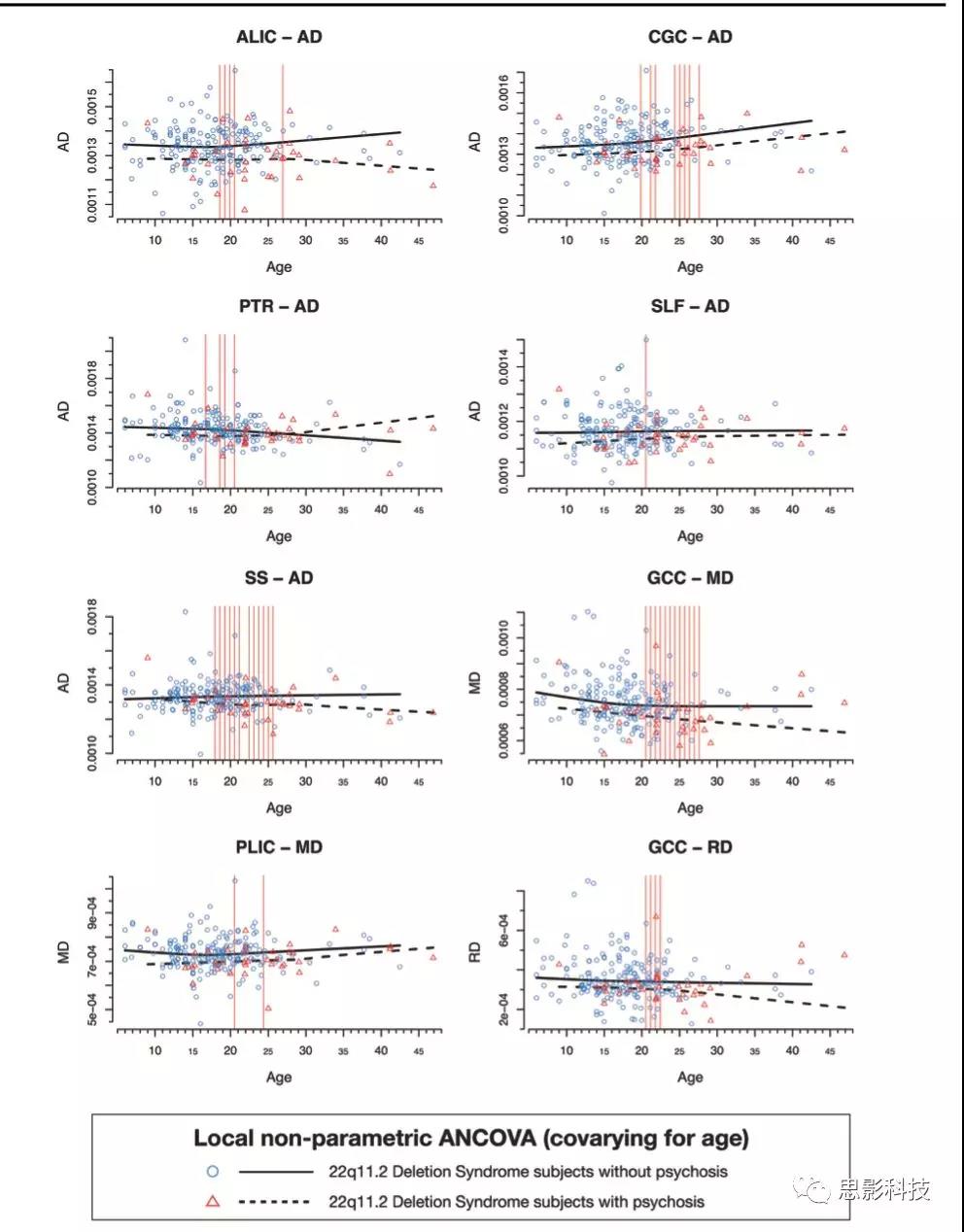
图3:患精神障碍与非精神障碍22q11DS非参ANCOVA比较
22q11DS精神障碍与特发性精分白质微结构的比较:
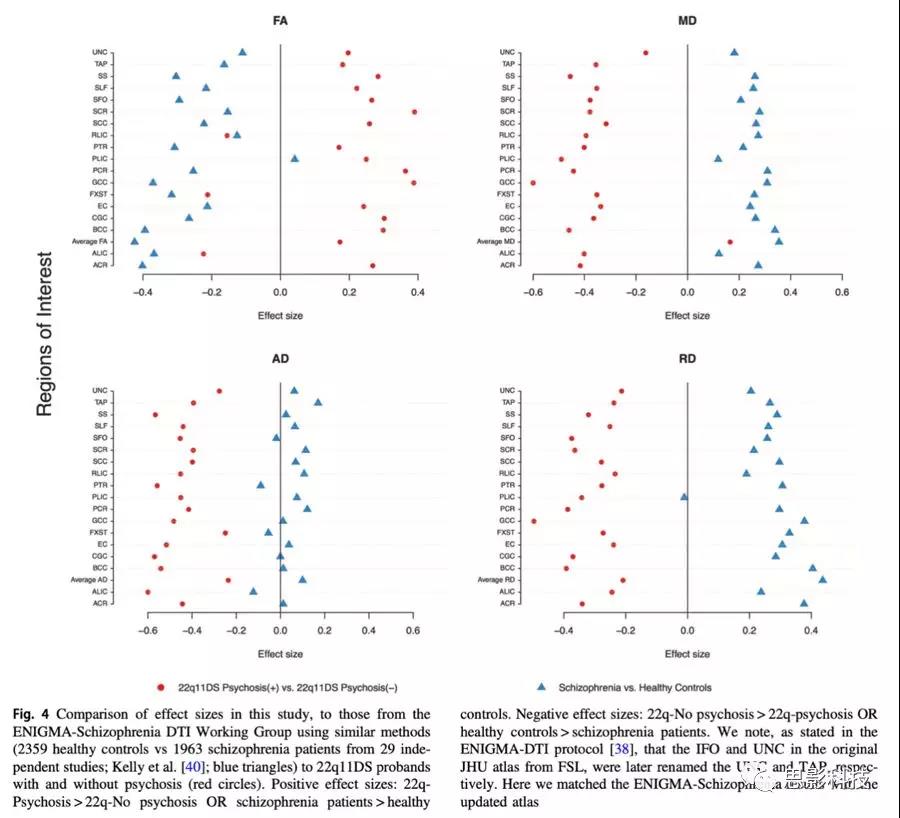
将22q11DS有精神障碍和无精神障碍的病例结果与精分病例对照结果(HC:2359,精分:1963)进行比较(图4)。22q11DS伴和不伴精神病患者之间与原发性精分相对于HC之间的效应比较有显著差异。具体来说,与未患精神障碍的22q11DS患者相比,22q11DS精神障碍者趋于较高的FA和较低的扩散值,而原发性精分患者较HC来讲表现出总体较低的FA和扩散值增加,尤其是MD和RD值。
图4:效应值比较
缺失型和IQ的影响:
染色体缺失的程度是否影响白质微结构?A-D大缺失的受试者在前放射冠和EC中显示出AD值降低的趋势,在TAP中显示出FA值升高;然而,经过多重比较校正后,与缺失大小相关差异并无统计学显著性。
此外,在HC组多个ROI中呈现出MD、RD和AD与IQ正相关,在TAP中呈现FA与IQ负相关。在22q11DS病例中,结果相似,但高IQ与PTR中的AD显著升高相关。在22 q11DS中,CC膝和SS中高AD与高IQ相关。虽然这些关系在经过多次比较校正后并不显著,但总体的研究结果表明,相对于典型发育对照组,22 q11DS的白质微结构和认知之间的关系需进一步探究。
本研究是迄今为止最大的一项在22q11DS中应用DTI探究白质微结构的研究。本研究路线允许跨中心对数据进行协调前瞻性Meta和mega分析,这与传统的结合文献中统计结果的Meta分析不同。本研究方法首次解决了由于样本量小和变量分析规程导致的低效能问题,这些问题导致了目前DTI研究的异质性和不清晰性。
与许多神经精神疾病的研究结果相反,本文研究结果显示,与HC相比,22q11DS的AD、RD和MD总体较低,FA存在区域性差异。较高的FA、较低的RD和AD(较低的MD)似乎是22q11DS主要白质纤维微结构改变的标志,尤其是连合纤维胼胝体。虽然这可能意味着髓鞘形成的程度更好,但在对本研究结果进行这一解释时必须谨慎,因为dMRI不能直接反映髓鞘形成的程度。正如在无髓鞘束中所显示的那样,各向异性不仅依赖于髓鞘的存在,而且对轴突密度也很敏感。RD对轴突密度和胞外间隙的大小敏感,AD对轴突直径和组织结构敏感。此外,由于轴突密度和髓鞘形成相关,因此,在解释人群间FA和RD差异时,不可能将两者分开。
本研究推测观察到的组间差异可能是由22q11DS中细胞膜累积周长的增加(归因于轴突组成、髓鞘形成和/或反应性星形胶质细胞的差异),这阻碍了垂直于白质纤维束的扩散,从而增加各向异性和降低RD。本研究发现22q11DS相对于对照组在连合纤维ROI(TAP、GCC、BCC和SCC)中FA值增高,在以投射纤维为主的ROI中没有发现差异(RLIC、SS、PTR和SFO),而与小鼠22q11DS模型一致,长联络纤维束(EC、SLF、FXST)的ROI中FA低。特别是,这项研究发现基底祖细胞的增殖被破坏,随后2/3层(而不是5/6层)的投射神经元的频率被改变。连合纤维和长连接纤维主要来源于投射神经元,即外层2/3的锥体神经元,而皮质纤维和丘脑皮质投射纤维则倾向于来源于皮层5/6的锥体细胞。此外,本研究表明,22 q11ds中胼胝体和长联络纤维间的白质中断性质可能不同,有必要采用先进的显微MRI技术来区分这些差异。因此,这些跨物种的发现共同表明了一个潜在的神经生物学模型,该模型中,22q11.2位点的单倍体不足导致大脑早期发育特定方面的损伤,及随后可能会增加22q11DS患者出现神经精神疾病风险的神经回路的改变。
本研究推测本研究结果可能与22q11DS患者白质的三种组织病理改变有关。首先,最近一项对一名3个月大的22q11DS婴儿的神经病理学研究报告表明,外层皮质层的神经元比率降低,深层皮质层的神经元比率增加。这与上述提到的22q11.2小鼠模型的发现密切相关。皮层2/3层的锥体神经元在相关联络脑区间产生大量的皮层-皮质轴突投射。这些轴突呈现在本研究中的大部分白质ROI中。因此,在22q11DS中,皮质联络区(皮层-皮质投射)之间的靶向-起源信号可能被破坏,影响轴突分化的必要信号启动,最终影响典型轴突直径分布的发育,从而改变白质纤维束中的RD和AD。此外,在22q11DS 中,PLIC是唯一显示较高AD值的ROI。在大鼠中,AD值与轴突直径改变和轴突弯曲有关。PLIC是本研究中唯一一个主要含有皮质纤维的ROI,其主要来源于皮质层5/6,这表明起源于较深皮质层的纤维束内的轴突大小分布可能与起源于外层的不同。对动物模型和人类死后脑组织的进一步研究可能会对阐明上述问题有所帮助。
其次,DTI指标异常也可能反映了继发于微血管损伤的胶质细胞变化。22q11DS成人尸检研究发现表明:两种深部白质胶质增生与脑血管改变有关。在小鼠脑损伤模型中,伴随脑对微血管损伤反应的胶质增生症与各向异性增加有关。第三,DTI指标可能受到早期发育过程中神经元迁移缺陷引起的白质异位神经元影响。这些在22q11DS患者的神经病理学和神经影像学研究中均有报道。尽管本研究没有发现任何异位,但细微显微镜下的异位仅能通过组织学检测。
FA、MD、RD和AD值的年龄轨迹,以及对照样本的峰值和最小年龄估计与之前报道的相似。然而,22q11DS患者的平均峰值FA和最小RD年龄都较大;相应地,在峰值和最小年龄后,FA和MD值的变化百分比也较小。如上所述,这些发现可能表明成熟延迟继发于深部白质轴突直径和组织的改变,这可能是延迟髓鞘形成过程的前兆。相反,正如22q11DS成人死后尸检报告显示,成熟后较小的百分比变化(由峰值FA和最小RD表示)可能表示白质的潜在组织变化,可能是由胶质细胞变化引起的。然而,尽管有统一的协议,但谨慎解释是值得的,因为各中心的年龄分布不同,且数据在较老的年龄范围内是相对稀疏的。
与一些单中心研究显示结果一致,22q11DS精神障碍症状严重程度与CC和长联络纤维的扩散率呈负相关,本研究发现在22q11DS精神障碍患者的CC膝的RD和MD较低,而长联络纤维(如SLF和CGC)的AD较低。有趣的是,在以皮层-丘脑和丘脑皮质纤维为主的ROI(ALIC、SS和PTR)中, AD显著降低。未来研究应前瞻性地探讨丘脑主要投射纤维束在22q11DS精神障碍症状发生和发展中的作用。
值得注意的是,22q11DS精神障碍患者的白质微结构改变与原发性精分患者的表现大不相同,主要涉及FA较高(而不是较低)和其他扩散系数较低(而不是较高)。先前对22q11DS和临床精神障碍高危青年的单中心研究也报道了这种相反模式。与皮层灰质的发现相反,22q11DS精神障碍患者表现出与原发性精分高度显著的重叠,表现为额颞区皮质明显变薄。因此,本研究结果表明,在22q11DS相关和原发性精分中,灰质和白质的神经解剖重叠模式明显不同,并提示不同的白质表型可能导致相似后续临床结果。
本研究在22q11DS中发现的AD改变,在精神障碍患者中更为严重,可能表明轴突直径改变和白质纤维束弯曲度增加。在皮层联络区之间的关键连接处有许多小而曲折的轴突,可能导致白质成熟度改变、结构失连接和可能的精神障碍。在原发性精分中,白质变性(脱髓鞘和较大直径轴突丢失)也可能导致轴突形态的破坏,同样导致皮质联络区之间的结构失连结。
本研究没有发现染色体缺失大小对白质结构的影响,也没有发现22q11DS和HC间白质微结构和IQ之间的关系存在差异。A-B缺失类型的样本量非常有限,并且各中心成像参数不同,这可能会影响研究结果。此外,在22q11DS患者中,给予高度可变的精神药物和共病治疗,其效果在这里无法进行系统的研究。先前,在一个包括许多与当前分析相同参与者的样本中,研究发现在22q11DS患者中,精神药物治疗与皮质厚度或皮质表面积没有显著相关性。此外,先前对原发性精分患者的研究发现,DTI检测到的白质改变并不是由抗精神病药物引起的。
通过多轴突采集和新生物物理模型,未来研究可能会解决轴突内外体积分数和直径对这些异常的贡献。定量磁共振成像转移和灌注成像采集可能有助于澄清22q11DS中的髓鞘异常或潜在的脑微血管病变。
总结:
本研究发现表明22q11.2缺失对白质微结构有很大影响。与HC相比,22q11DS的AD、RD和MD总体较低,FA存在区域性差异。较高的FA、较低的RD和AD(较低的MD)似乎是22q11DS主要白质纤维微结构改变的标志,尤其是连合纤维胼胝体。同时,在测试年龄、染色体缺失、IQ以及精神疾病与白质微观结构受损关系的分析中发现,年龄并未表现出明显的发育效应,而染色体缺失类型和IQ表现与白质损伤没有显著关联。对患精神疾病的22q11DS患者的研究则表明,22q11DS精神障碍患者的白质微结构改变与原发性精分患者的表现大不相同,主要表现为FA较高(而不是较低)和其他扩散系数较低。
此外,在动物模型中,22q11DS中主要皮层-皮质和皮质-丘脑/丘脑-皮质连接的中断可能归因于早期神经发生的中断。未来研究将帮助确定这些改变的神经生物学基础。
如需原文及补充材料请加微信:siyingyxf 或者18983979082获取,如对思影课程感兴趣也可加此微信号咨询。

微信扫码或者长按选择识别关注思影
非常感谢转发支持与推荐
欢迎浏览思影的数据处理课程以及数据处理业务介绍。(请直接点击下文文字即可浏览,欢迎报名与咨询):
第九届磁共振脑影像结构班(南京)
第六届任务态fMRI专题班(重庆)
第十八届脑电数据处理中级班(南京)
第十九届脑电数据处理中级班(南京)
招聘:脑影像数据处理工程师(重庆&南京)