

偏头痛是世界范围内发病普遍、致残率高的疾病之一,产生巨大的社会经济负担。然而,偏头痛的发病机制不明,尚无可识别病理改变的诊断标志物。针对偏头痛的特异性标志物将有助于诊断及发病机制的研究,对开发新疗法存在潜在的临床价值。在过去数年中,神经影像与遗传学的研究对于标志物的识别带来了重要进展,越来越多的脑成像研究为研究偏头痛发作期及发作间期临床症状的潜在脑机制提供了重要信息。同样,大规模的全基因组关联分析已识别与先兆偏头痛与无先兆偏头痛这一常见偏头痛形式相关的遗传变异。共44个独立的单核苷酸多态性位点被发现与偏头痛的发病机制存在密切相关,并为血管机制的参与提供了新证据。神经影像与遗传作为偏头痛标志物具有极大潜力。本文对现有及潜在的神经影像与遗传标志物结果进行总结,并对标志物对于机制的理解及潜在的神经影像及遗传联合进行讨论。本文由澳大利亚学者发表在NATURE REVIEWS NEUROLOGY杂志。
正文:
偏头痛发病普遍,在2015年全球疾病负担研究中,偏头痛因造成长期残疾在全球310种疾病中排名第7。偏头痛所造成的经济负担在所有中枢神经系统疾病中排名第2,仅在欧盟国家年经济负担达到1110亿英镑。
根据国际头痛学会第二版诊断标准,也是目前研究中最为常用的诊断标准,偏头痛可分为两种常见类型:无先兆偏头痛(MO)及典型先兆偏头痛(MA)。这两种类型占据偏头痛的90-95%,余下5-10%为少见的伴有特异性神经系统症状的偏头痛。偏头痛确切的病因及不同类型之间病理生理差异目前尚不明确,缺乏具有诊断价值的特异性病理改变。因而临床对于偏头痛的诊断主要依据头痛症状的特征及相伴随的症状,其内科系统、神经系统查体及实验室检查通常正常。提高对偏头痛病理生理机制的理解对偏头痛的临床诊治及发展潜在的有效治疗靶点提供巨大帮助。
挖掘影像及遗传数据
偏头痛的发作常伴随非疼痛症状,包括:感觉过敏、恶心、认知障碍及情绪异常,表明偏头痛会影响整个大脑。影像对于理解大脑机制提供潜在研究价值。同样,40-60%偏头痛的遗传率表明遗传因素在疾病中发挥重要作用,识别与偏头痛相关的特异性基因变异对于探索疾病病因具有帮助。其他研究方法也可提供对疾病的见解,包括:生物液体分析(血清、唾液、脑脊液),神经生理测评分析(诱发电位),脑磁图与脑电图。但目前针对偏头痛,进展迅速且最具稳健性及信息量较大的生物标志物研究仍为影像学与遗传学研究。
偏头痛的影像学研究
偏头痛存在多种临床表型,很多方面表现不同,包括:发作频率(发作性或是慢性,慢性包含一个动态的慢性化过程)、有无先兆、感觉、情绪、运动及自主神经系统改变;历经发作前期、发作期、发作后期及发作间期;先兆症状,可出现在发作前2天,包括疲劳、认知、情绪改变或颈强直;性别相关差异。多数改变反映大脑状态的变化(感觉、情绪、自主神经及认知);因此,通过神经影像将临床表现转化为大脑表型,评估大脑功能、结构、代谢是具有意义的(Fig1)。
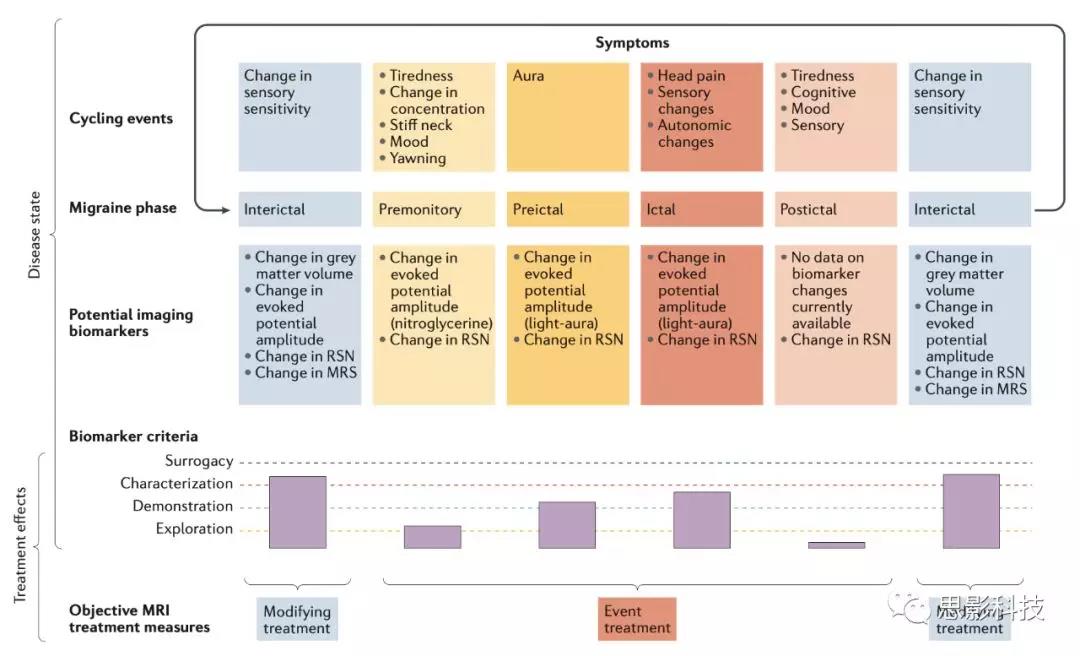
图1.偏头痛状态进展的标志物上两列为疾病状态及潜在的影像标志物。“Biomarker criteria”一列总结每个疾病阶段当前标志物研究进展---探索、验证、描述特征或是替代。标志物可被用来评估药物疗效或疾病状态,如:慢性化、改善或症状治疗。
神经影像存在的问题在于难以区分所观察到的改变是致病性、疾病所造成的,或是兼而有之。随着年龄增长或偏头痛发作频率增加,大脑存在进行性改变表明疾病驱动大脑变化。相反,脑改变的神经影像标志物看似是可逆的,至少会随着发作的减轻而变化表明标志物可表征疾病活动。然而,一些变化所代表的生理意义并不清楚,例如:在药物使用过度例如阿片类药物引起的头痛中,当药物撤退会观察到一些脑区变化,但其中因果关系未知。且这些变化在其他疼痛中是否存在尚未可知。
总之,神经影像可作为疼痛进展的标志物,且在偏头痛领域从功能、结构、代谢方面的影像研究呈指数增长。在接下来的部分,将着重评估神经影像包括功能磁共振(fMRI)、PET用于识别偏头痛及其进展的标志物的开发及应用。
动态偏头痛大脑成像
偏头痛大脑成像多数在发作间期进行,发作期较少,发作前期、先兆期及发作后期的研究更少。尽管如此,偏头痛发作期的神经影像研究依然为围发作期的各个方面提供了新思路。这些观察有助于对偏头痛及发作前、中、后期的潜在机制的理解(图2)。

图2. MRI测量的定义及偏头痛脑功能及结构示例血氧水平依赖性(BOLD)功能磁共振成像(fMRI)显示丘脑激活伴痛觉异常。同一患者的强迫症状态(偏头痛期间对热刺激的敏感性增加)和基线状态之间的BOLD-fMRI信号差异显示在红橙色区域。静息状态网络(RSN)fMRI显示顶额叶静息状态的具体变化。体素形态计量学(VBM)显示中央后回的区域皮层厚度增加,高频偏头痛组相对对照组皮层厚度增加。扩散张量成像(DTI)显示偏头痛患儿与健康对照组的白质扩散率显著降低(P<0.05,FEW校正),在MNI空间的分数各向异性(FA)模板上可视化。磁共振波谱(MRS)结果显示了偏头痛患者MRS体素记录的2D J空间1H MRS数据成像,显示兴奋或抑制通道的改变。如果您对以上脑影像数据处理方法感兴趣,请直接点击下文思影科技课程,或添加微信号siyingyxf或19962074063进行咨询:
第二十八届磁共振脑影像基础班(重庆)
第十四届磁共振脑网络数据处理班(重庆)
第九届磁共振脑影像结构班(南京)
发作前期成像:
识别围发作期标志物用来预测发作期尤其重要,因为发作早期治疗敏感性更高。将这些标志物转化为临床实践,有助于通过治疗稳定偏头痛的发作阈值,预防进展至发作期。
前兆期:
目前尚无神经影像研究捕捉到一个自发性偏头痛的先兆期的脑改变,尽管先兆症状出现在7-88%偏头痛中。在一例偏头痛患者的研究中,对患者施加疼痛刺激(通过聚四氟乙烯管向鼻腔输入气态氨)后进行影像扫描,持续30天,发现围发作期下丘脑功能连接发生改变。进一步研究发现偏头痛患者存在下丘脑功能异常,包括:下丘脑对不同波长的光的反应性发生改变。偏头痛患者被认为存在潜在的自主神经系统异常,加之下丘脑的证据,及由PET研究所支持的其可引起的自主神经系统的异常均表明自发性偏头痛发作后期下丘脑存在受累。
在体神经影像研究中,使用硝酸甘油诱发偏头痛发作,发现后外侧下丘脑、脑干(包括中脑被盖区、中脑导水管周围灰质及脑桥背侧)及部分皮质区域(包括枕叶、颞叶、前额叶)在先兆期存在激活。在同样的患者进行同样的硝酸甘油刺激下,发现光过敏现象,表明视觉皮层的超敏性。虽然硝酸甘油的影响是否为症状的一部分并不清楚,但这类研究有助于理解先兆期的神经基础。
先兆期与皮层扩散性抑制:
2001年一项开创性影像研究发现了在视觉先兆期间视觉皮层的激活。该观察揭示了一种类似于皮层扩散抑制(CSD)的自发性电生理事件存在于人类偏头痛患者中,且这种电活动事件在小鼠中可激活脑膜痛觉感受器。这些研究表明,理解先兆期相关的广泛大脑改变的潜在意义及伴随CSD出现的亚临床改变的可能性。例如:如动物实验所验证的,在人类中CSD出现于皮层及皮层下结构,或可成为以CSD为靶点或限制其敏感性的特异性治疗方法。正如在偏头痛的大鼠模型中CSD样现象出现在皮层及皮层下结构,通过fMRI,在典型先兆偏头痛(MA)患者相较MO(无先兆偏头痛)患者中,已发现脑功能存在差异,这些差异包括半球间差异(具有固定侧别的先兆患者)、发作间期视觉皮层对于视觉刺激的高反应。
三叉神经系统改变:
三叉神经系统是具有特异性且被广泛研究的神经解剖通路,包含三叉神经节,可通过fMRI进行评估。在一项偏头痛患者的fMRI研究中,随着痛觉刺激,三叉神经节激活增加,表明处于偏头痛发作的围发作期。提出偏头痛发作易感性相关的潜在过程,可能基于与多种因素相关的稳态,包括睡眠、应激、炎症等。
发作期成像
发作期改变:
MRI较难捕捉偏头痛整个发作期脑功能改变,因为偏头痛自发发作且较难诱发。目前尚无针对偏头痛自发性发作的功能成像研究,有部分研究捕捉到正处于发作中的患者。识别发作期影像标志物,有助于确定受影响且在未来发作中较为敏感的脑区。丘脑(后部)BOLD信号与皮肤触觉诱发痛存在关联,并由动物实验证实。颞叶过度激活也为理解偏头痛复杂行为的机制提供思路。同时在发作期观察到下丘脑与三叉神经核团对伤害刺激反应过程耦合改变,表明下丘脑与脑干功能连接的改变对于驱动发作十分重要。最后,μ阿片类受体在发作期间发生动态变化,为理解慢性化过程提供了见解。尤其是疼痛调节区域受体数量的改变(如中脑),参与触觉诱发痛生理表现。
脑干改变:
多个脑干核团参与自主神经功能,例如,孤束核、延髓最后区(恶心与呕吐中枢);迷走神经核团,包括副交感神经及参与应激反应的蓝斑核。这些脑干核团均与偏头痛相关,研究发现硝酸甘油诱发偏头痛脑干激活。脑干核团的MRI研究仍存在技术瓶颈,多数偏头痛研究尚未得到详细的神经解剖学信息及这些核团与行为相关的特异性活动。
发作期脑干与其他脑区功能连接发生特异性改变(如蓝斑核及下丘脑),下丘脑背侧-喙侧脑桥连接支持蓝斑核在偏头痛中发挥作用。在偏头痛发作中观察到脑干对伤害性刺激的偏侧激活(中脑、脑桥腹侧、脑桥背侧以及三叉神经脊束核)。同时也观察到脑干疼痛调节区域功能连接发生改变。脑干可能是关注疾病变化的潜在标志物区,其多个核团参与偏头痛的主要病理改变,包括:疼痛调节(通过中脑导水管周围灰质及中缝核)、自主神经改变(通过迷走神经核团)、大脑过度兴奋(蓝斑核),针对脑干标志物的研发需克服其技术瓶颈。
发作后期成像
偏头痛发作后会表现出显著的困倦及认知情绪改变,但目前尚无相关神经影像研究。仅有一项研究评估一例女性患者偏头痛发作的整个过程(发作前、中、后期)BOLD信号改变,其中发作后期视觉皮层BOLD信号发生改变,与发作间期相比,发作后期视觉皮层对伤害性鼻刺激反应活动增加,表明存在光过敏。
发作期与发作间期成像
了解同一患者大脑发作期及发作间期变化有助于理解大脑动态反应的机制,部分研究评估患者发作期及发作间期状态。其中一项研究观察到发作期及发作间期患者枕叶激活降低。在一项6例被试的PET研究中,发作期与发作间期相比较,
发作期多个脑区激活增加:
桥脑背侧、小脑、丘脑、前扣带、后扣带、前额叶、岛叶及颞叶。另一项研究中,颞叶对热痛刺激反应在发作期及间期较正常人发生改变,表明这些脑区在偏头痛患者中存在过度兴奋。
“静态”偏头痛大脑成像
目前多数关于偏头痛脑成像研究多关注发作间期,并发现较健康对照相比,发作性或慢性偏头痛患者存在脑功能、结构及代谢改变。这些改变是发病前已出现或是疾病导致的结果并不清楚,现有数据趋向于认为偏头痛发作参与改变。支持这一观点的证据包括但不限于发作频率差异、慢性化逆转、性别差异、围青春期改变。MRI评估发作间期脑形态学、功能、代谢改变,发现多种疾病的重要标志物。
脑体积:
研究发现已发现偏头痛患者存在多个皮层及皮层下(小脑、基底节、导水管周围灰质)灰质体积的显著改变(增加或萎缩)。树突结构复杂性改变参与这一变化。脑体积的改变可能与偏头痛患者行为存在相关(例如:额中回与额下回体积改变与认知功能)。研究也刻画偏头痛患者白质损伤,但其病理机制及对脑功能所造成的影响并不清楚。在MA患者中,白质损伤显著增多且与疾病严重程度相关,而因果关系并不清楚。与偏头痛相关的主要事件包括卒中,改变大脑结构与功能。目前存在的问题为作为标志物识别大脑随时间的改变,灰质体积萎缩可随治疗逆转,或与发作频率、致残、性别或年龄相关。
S1皮层与偏头痛:
在发作性偏头痛研究中发现,躯体感觉皮层(S1)及位于感觉区域中代表头部部分皮层的厚度增加。后者伴随应对疼痛刺激时功能连接改变。同样在动脉自旋标志物测量脑血流及体感诱发电位研究中发现发作间期S1区域兴奋性增加。这些变化可能参与丘脑-皮层调节感觉接收信息增加。因此,三叉神经-丘脑-S1系统的状态可能预测治疗有效性。
情绪状态:
偏头痛对脑功能最有趣的改变在于对大脑复杂处理过程的改变,如:情绪。情绪处理脑区包括基底节、前脑岛、杏仁核、前扣带皮层,其结构、静息态活动发生改变。除此之外,对于厌恶性刺激的反应偏头痛患者较正常人存在不同。较少有研究评估偏头痛情绪处理,但是在疼痛领域,情绪处理的改变可以预测疼痛的慢性化。这些发现可改变我们对偏头痛的认知,为治疗方案的选择提供参考。
化学失衡:
应用磁共振波谱(MRS)为认识疾病提供新的信息,如:测量某些皮质兴奋性神经递质(N-甲基-d-天门冬氨酸,NMDA)与抑制性神经递质(GABA),鉴别偏头痛患者与健康对照。研究发现在前扣带皮层和脑岛,偏头痛患者兴奋性氨基酸N-乙酰天冬氨酰谷氨酸及谷氨酰胺存在显著异常。MRS为评估神经递质及相关治疗提供技术支持,未来对于预测疾病治疗的反应性具有一定意义。
内源性疼痛调节:
头痛严重程度的增加、痛觉超敏及头痛慢性化为偏头痛的特性之一,表明内源性疼痛调节发生潜在改变。研究发现偏头痛患者存在下行性调节通路异常,即抑制系统不能有效阻止疼痛。增加下行性抑制作用可能成为治疗偏头痛的潜在方法,这类研究相关标志物可有助于筛选适用于该类治疗的患者。
过渡状态的成像
研究本文定义的过度状态包括:反映生理状态(性别、青春期)及疾病状态的恶化与改善(发作频率与发作强度的改变)。
偏头痛慢性化:偏头痛慢性化与多种危险因素相关,包括:基因、环境因素、并发症(压力、肥胖)、皮肤异常性疼痛、心理异常(抑郁、焦虑)、睡眠障碍、高偏头痛发作频率及药物的过度使用(包括阿片类药物和曲坦类药物)。由发作性偏头痛转变为慢性偏头痛是一个动态过程,慢性偏头痛的影像研究对于帮助理解疾病进展的潜在机制提供帮助。
目前,对于偏头痛慢性化的生物学研究较少,但脑结构及功能改变与偏头痛发作频率增加相关。PET研究目前已发现慢性偏头痛患者存在脑干代谢增加。慢性偏头痛MO患者眶额部皮层及左侧枕中回灰质体积下降,且眶额皮层灰质体积有助于预测对于药物过度治疗反应。在一项23例小样本慢性偏头痛的研究中,对于逆转慢性偏头痛治疗存在反应的患者较无反应显示出初级躯体感觉皮层、前脑岛、左侧颞上回、左侧岛盖部皮质增厚,同时疾病病程与额顶、颞枕区域皮层厚度呈负相关。这类针对慢性期、发作期治疗疗效的研究可提供预测影像标志物,表明疾病对大脑的调节作用。
青春期及性别:
偏头痛与性别的关系不仅表现为发病率的差异(women 7:1 over men),同时灰质体积与功能存在差异。同样的,青春期男性与女性偏头痛患者也存在脑功能与结构差异,而这些差异在青春期之前基本不存在。因此,除了遗传倾向,荷尔蒙水平及其他因素(药物影响包括抗偏头痛药物及节育治疗)的研究有助于理解偏头痛的易感性及疾病进展。
治疗影响:
采用神经成像评估新型偏头痛药物疗效具有临床意义,有助于理解药物作用机制,研发预测药物疗效及副作用的有效偏头痛特异性脑影像标志物。较少有研究评估抗偏头痛药物在健康人群及患者中对于脑功能及结构的影响。已有研究报道通过fMRI及PET评估抗偏头痛药物(如曲坦类)对前临床模型及1次偏头痛发作的健康志愿者的影响。在前临床动物模型中,曲坦类药物的慢性化治疗使大脑状态增敏,在应激状态下(暴露于光照下)产生CSD样现象。同时,药物对多个静息态脑网络产生显著影响,包括:默认网络、自主神经系统、基底节、凸显网络及感觉运动网络。在健康被试中,舒马曲坦治疗显著增加三叉神经核团及丘脑活动,同时抑制丘脑-皮质连接。曲坦类药物逆转偏头痛患者头痛症状,但下丘脑、脑桥及中脑在自发性偏头痛发作中依然被激活。这些研究对于发展评估药物疗效的影像标志物或治疗靶点提供潜在参考。
偏头痛的遗传标志物
单基因遗传
早期对于偏头痛遗传机制的研究是通过家族性偏瘫型偏头痛(FHM)进行的,这是一种罕见的符合孟德尔遗传定律的具有先兆症状的偏头痛。目前存在3种FHM相关的基因突变:FHM1型,CACNA1A错义突变,编码电压门控钙通道α亚单位;FHM2型是ATP1A2突变,编码ATP酶Na+/K+交换泵亚单位α2;FHM3型是SCN1A突变,编码电压门控钠通道α1亚单位。这三种FHM相关基因均参与离子通道转运,细胞及动物实验表明,神经元超兴奋是FHM头痛及先兆症状的一种潜在细胞机制。FHM的主要症状头痛、先兆症状伴随恶心、畏光、声音恐惧症与先兆偏头痛症状类似,表明FHM相关基因或编码其他金属离子通道基因参与MO及MA易感性。
多基因遗传
相对于FHM,多数偏头痛并非单个高影响力基因突变所致,MA、MO均为多种基因、环境及其相互作用的复杂疾病。目前有两种方法用于识别MA、MO特异性基因变异:候选基因关联研究(CGAS)及全基因组关联研究(GWAS)。CGAS涉及对候选基因变异频率在偏头痛患者和一般人群或非偏头痛人群的比较。CGAS由于设计简单且成本较低成为研究偏头痛遗传变异的主要方法。候选基因的选择是基于基因已知或预测生物学功能及其与偏头痛病理机制理论的相关性。然而,CGAS一般在小样本(100-200病例对照组)对少数变异进行检测,缺乏稳健性,导致假阴性及假阳性率较高。本文对此不进行综述。由于对大样本大量基因多态性分析的需求,成立国际头痛遗传学联盟。对基因分析技术的改善使得IHGC进行大规模关联分析确定常见偏头痛的危险基因。首批IHGC研究分析841例患者及884例健康对照的155个编码离子通道基因的5257个单核苷酸多态性(SNPs)。SNP均经过精心挑选,确保出现所有的等位基因(次等位基因频率MAF>0.01)并通过微阵列分型。对于所出现的最强信号的SNPs在4组独立病例对照样本进行验证,共包含2835例MA或MA患者及2740例对照。该研究发现所检测的离子通道基因常见变异(MAF>0.1)具有中等效应量(基因型相对风险1.40-1.65),不影响常见偏头痛的易感性。
GWAS(全基因组关联研究)
基因分型阵列技术的发展,可进行超过300,000 SNPs测序分型,使得大规模GAWS成为可能。GWAS可检测基因组中大多数常见的SNP变异,结合大量遗传数据,严格的统计,使得所得SNP危险位点具有稳健性,误差较少。
基于临床中心研究:
最早两项IHGC GWAS病例来自专科欧洲头痛临床中心。首项2010年IHGC GWAS包含2731例MA患者及10747例匹配的对照。研究确定来自8q22.1染色体的SNP(rs1835740)基因,编码异粘蛋白(MTDH)及羧肽酶Q(CPQ)。该关联基因在3203例仅MA患者,MO患者或MA及MO患者与40062例对照组进行了验证。研究rs1835740基因型如何影响临近基因的表达发现rs1835740是MTDH基因的顺式调节元件。有趣的是2011年一项欧洲GWAS研究中未发现达到显著性的位点,但是先前鉴定候选基因中MTDH与偏头痛显著相关性(Bonferroni-adjusted gene-based P = 0.026)。
2012年IHCGGWAS研究较2010年样本量减少,确定6个MO患者SNP危险位点:MEF2D基因rs3790455 (肌细胞增强因子2D), TGFBR2 附近rs7640543 (转化生长因子β受体2), PHACTR1基因rs9349379(磷酸酶和肌动蛋白调节器1), ASTN2基因rs6478241(星形肌动蛋白2), TRPM8基因rs10166942(瞬时电位阳离子通道受体亚家族M成员8),LRP1基因rs11172113(LDL受体相关蛋白1)。TRPM8及LRP1基因SNP,及PRDM16基因rs2651899在另一项女性大样本偏头痛研究中被验证。
基于人群的研究:
2011年IHCGGWAS研究5122例女性偏头痛患者较18108例女性对照的基因变异,并确定了7个SNPs。结合验证队列,得到PRDM16, TRPM8 及LRP1 位点与偏头痛的相关性。随后对前述3项GWAS研究及其他研究进行Meta分析得到12个与偏头痛相关位点。除新发现的5个SNP位点,对前述2010年IHCG GWAS研究MA相关位点得到验证,除rs1835740 SNP(MTDH附近)。对于146项关联度最强的SNP指标的分析发现效应量在MA及MO亚群之间、基于临床中心及基于人群之间、男性与女性之间相似。由于146个SNP是通过固定效应meta分析确定的,该模型假设在所有研究具有同等效应,因此结果是可以接受的。但是对于超过23000的SNPs,效应一致性分析对整体遗传重叠的评估发现,主要亚组遗传风险的一致性并不限于全基因组显著性及已被鉴定的SNP。因此,不同亚组间遗传风险的差异被相似性所掩盖,其他偏头痛GWAS数据集的meta分析,不考虑其亚组构成,可有助于确定新的易感位点。
因此,IHGC进行第4次GWAS研究并发表于2016年,数据来自59674例患者及316078例对照。该研究确定了与偏头痛相关的38个基因组区域包含44个独立的SNPs位点。3个SNP位点具有全基因显著性(rs1835740 near MTDH115, rs10915437 near AJAP1 and rs10504861near MMP1)。这些SNP位点是分别通过与偏头痛各亚型的相关性确定的(MA、临床MO或MO及MA)。
因此,目前尚需要在同质化组内最终确定这些位点的稳健性。2016IHCG GWAS对800万个常见SNPs(MAF>0.01)基因型进行分析,提供了一个对常见SNP变异的更全面的评估,其与偏头痛风险相关的变异及更准确的关联信号定位。除此之外,计算分析SNP可信致病集,对于认识偏头痛的病理生理机制提供帮助。一般来说, 在GWAS鉴定的基因组区域的连锁不平衡导致难以确定可能的致病SNP及变异可能的生物学功能。以往偏头痛GWAS研究讨论相关SNP可能的生物学效应是通过最为显著相关的位点所在的基因或SNP最接近的基因。
然而,基因对偏头痛的生物学作用除物理位置以外的证据较少。2016IHCGGWAS采用贝叶斯方法,包含关联分析及连锁不平衡结构,确定得到一个99%概率包含致病性SNP的可信SNP集。可信SNP的位置被用来确定与偏头痛最有可能相关的基因。除此之外,采用DEPICT(数据驱动,对复杂特性的优先级集成的表达)方法确定每个GWAS位点的致病基因。这种方法对预测功能与其他相关位点的基因共享频率比期待较高的基因进行优先排序,得到基因列表(Table1)。可信SNPs分析及DEPICT的基因优化得到37个基因可能是致病基因,其中27个相关区域各一个基因,5个相关区域2个基因。
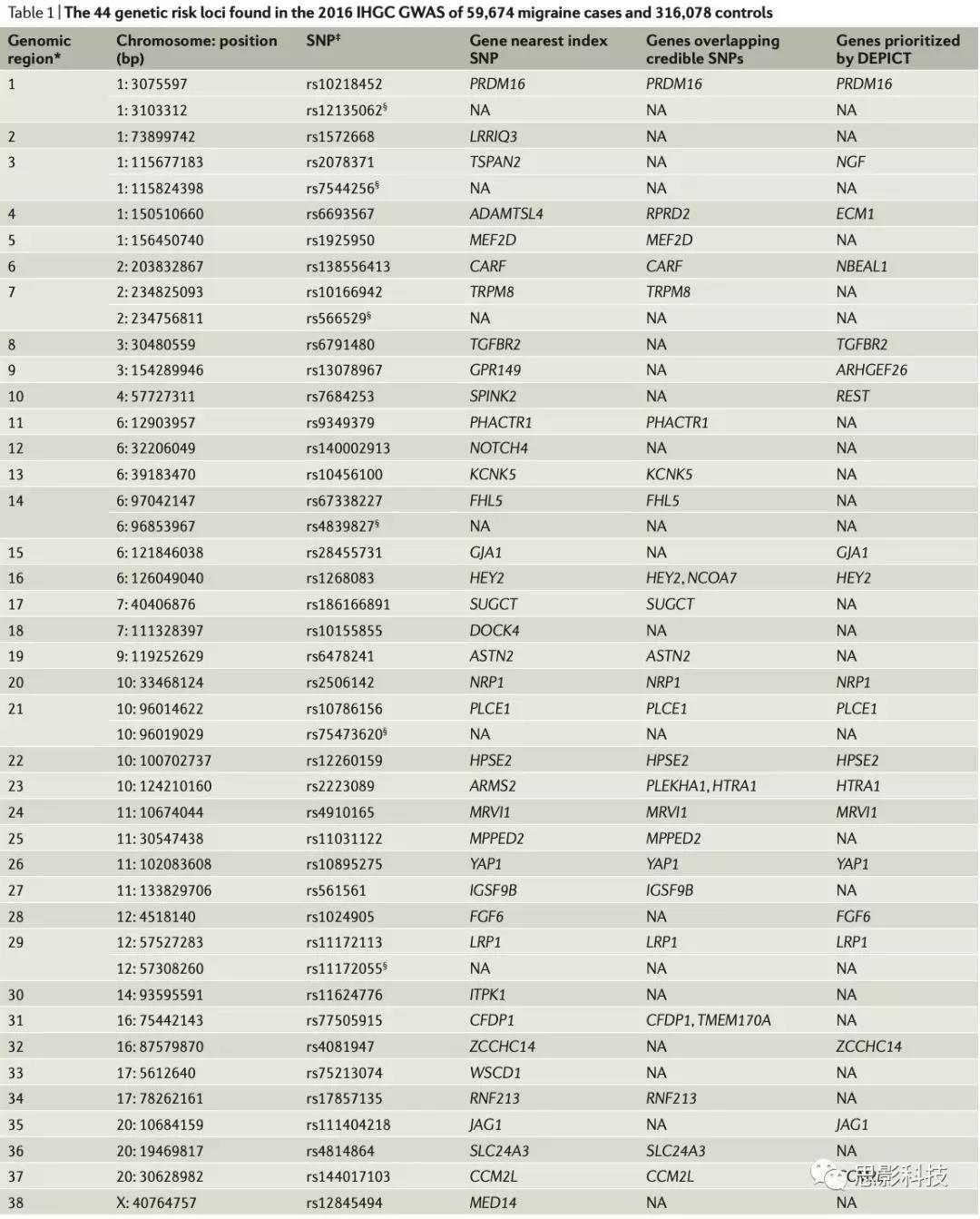
表1. 2016年IHGC GWAS中发现的44个遗传风险基因位点
可能的致病基因
为了描述37种可能的致病基因(表2)的特征,有研究应用g:GOSt工具,一种基于网络的基因功能谱分析软件g:Profiler128 (http://biit.cs.ut.ee/gprofiler/),应用GeneOntology(GO), Kyoto Encyclopedia of Genes and Genomes (KEGG)及Reactome数据库进行富集分析,高度可靠,人工选择人类基因的功能注释(图3)。
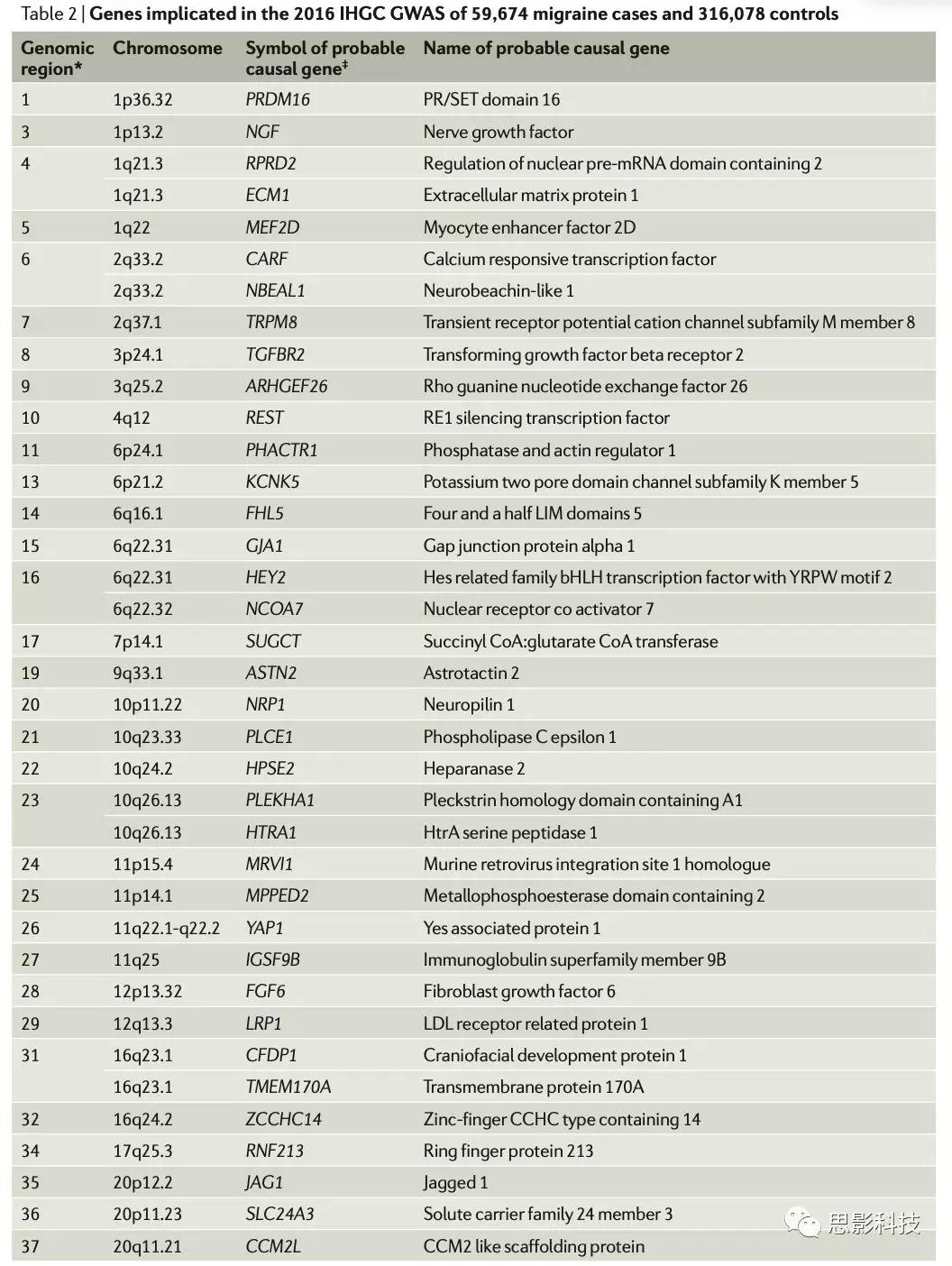
表2.Genesimplicated in the 2016 IHGC GWAS of 59,674 migraine cases and 316,078 controls

图3.37中偏头痛相关基因GO、KEGG、Reactome通路富集词云图
对于37种基因,富集出102种通路。其中血管相关通路明显富集。实际上,9种基因与“循环系统发育”相关。相反,仅有两个金属离子通道通路富集,分别为“钾离子跨膜转运体活性”(GO:0015079; REST, KCNK5, and SLC24A3)及“响应钙离子的RNA聚合酶II启动子转录的正调控”(GO: 0061400;CARF)。
通过生物信息学方法研究纳入的GO功能注释为血管相关通路富集提供更多证据,添加4种基因(CCM2L、FGF6、GJA1、YAP1)将“循环系统发展”基因注释数量提高至13。有趣的是,这种分析也强调了5种新的金属离子相关通路:“细胞钙离子稳态”、“钙离子稳态”、“金属离子跨膜转运体活性”、“离子通道活性”及“钾离子运输”。扩展通路分析至所有人类基因,进一步强调血管相关通路,例如:“循环系统发育”及“脉管系统发育”。
实际上,共有25种血管相关通路在15种血管相关基因中高度富集。最受关注的金属离子相关通路是“金属离子结合”,富集在11种金属离子相关基因中。这些结果表明离子通道功能紊乱可能并非为偏头疼发病机制的主要部分,但基因引起的金属离子稳态变化可能会促成偏头疼易感性。组织表达富集分析同样在2016IHGC GWAS中进行:在50kb以内的可信SNPs基因集表达在42个不同的组织中。研究结果表明,这些与偏头痛相关的基因在动脉及胃肠组织中显著富集。
进一步研究将分析的组织分为4组:脑、血管、胃肠及其他,8种基因在血管组织中显著富集。上述通路及组织表达富集分析的研究结果为偏头疼易感性的血管功能障碍提供强力的证据。这个结论对于存在争议的偏头疼是否为原发血管功能障碍或大脑异常继发血管改变十分重要。并且,偏头痛的血管机制可以解释其与卒中及心血管疾病并发以及遗传的重叠。实际上,9种与偏头疼相关的基因与血管疾病均有相关性。一个特殊的发现是位于17q25的RNF213蛋白的伴有烟雾病的基因突变,此病临床特点具有偏头疼样头痛发作的特点。
与其他表型关联
一些与偏头疼相关的基因位点也可能与其他表型相关。例如,偏头痛相关SNP rs11624776位于ITPK1附近,也是与甲状腺激素减低相关的SNP。甲状腺功能减低与头痛之间的相关性研究最早自1948年,相较于发作性偏头疼患者,甲状腺功能减低在慢性偏头疼患者中的发病率更高;并且患有偏头疼的患者其将来患有甲状腺功能减低的可能性更大。
偏头痛相关的SNP rs9349379位于PHACTR1基因,与冠心病、冠状动脉钙化及颈动脉夹层等相关;与SNP rs9349379 存在连锁不平衡的SNP与早发性心肌梗塞(rs12526453)、冠脉疾病和大动脉缺血性脑卒中(rs4714955)相关。偏头疼相关SNP rs11172113位于基因LRP1,其与肺功能及颈动脉夹层相关,与rs11172113连锁不平衡的SNP rs1466535与腹主动脉瘤相关。
与附加偏头痛位点上存在连锁不平衡的SNPs可能与其他表型相关。SNP rs10789369与偏头痛相关rs1572668存在连锁不平衡,位于LRRIQ3附近与精神分裂症相关。另一SNP(rs2341263)与rs1572668存在连锁不平衡,与血清代谢物X-11787水平相关,后者为急性心脏事件的危险因素。与rs1268083存在连锁不平衡的SNP rs9388451位于HEY2附近的与Brugada综合征相关,后者为一种致死性心律失常疾病;与rs10166942存在连锁不平衡的两种SNPs位于TRPM8附近(rs10187654及rs7577262)与肺功能减低及冷刺激试验下血压反应性相关。SNP rs7940646与rs4910165存在连锁不平衡,位于MRVI1基因,与血小板聚集相关。与rs138556413存在连锁不平衡的SNP rs6725887位于CARF基因,与早发性心肌梗塞、冠心病及冠状动脉疾病相关。与rs138556413存在连锁不平衡的SNP rs7582720与冠脉疾病及大动脉缺血性卒中相关。与rs6478241存在连锁不平衡的SNP rs7852872位于ASTN2基因,与海马体积相关;
值得注意的是,因为此该SNP位于编码星形肌动蛋白2的基因内含子上,星形肌动蛋白2为一种表达在神经元上的细胞黏合分子,被认为在胶质介导的神经元迁移中发挥作用,在ASTN2位点的基因突变可能会导致偏头疼患者脑结构及功能异常。虽然鉴别偏头痛相关风险因子与其他表型的相关可能提供新的偏头疼并发症的生理机制,单个偏头疼变异仅增加很小的相关并发症风险。如表型本就罕见(如烟雾病),那这些相关性仅存在于比例很少的人群中。
基因相关性作为偏头疼标志物
在过去的几年,证实与偏头疼相关的风险基因位点显著增加。随后有研究致力于发现致病基因,研究发现多种不同通路异常,包括血管通路、电解质稳态及神经元迁移等等。基于以上研究结果,未来的研究需致力于全面阐明偏头疼患者每个基因位点的功能影响,区别该位点是否具有诊断性或预测性价值,为偏头疼患者提供个体化治疗方案。为了达到此目的,大样本量、根据不同表现细分的临床表型群组需在未来的研究中得到应用,以达到明确每个位点独立的作用及其相互之间的作用。
结合影像学及基因研究
生物遗传标志物的可能性
过去几年中针对偏头痛的影像学研究或遗传方面的研究已有许多发现,如可以将二者结合则可能对偏头疼的发病机制有更进一步的探索(图4)。
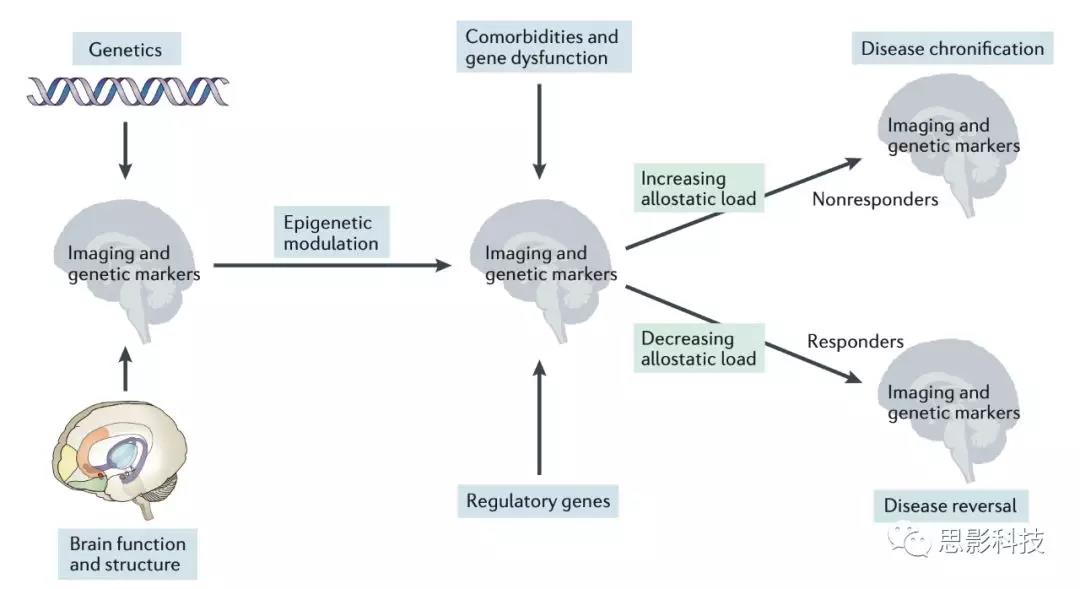
图4. 整合影像学与遗传学偏头痛标志物
发现一种新的生物标志物需以下步骤:研究探索,发现可能的潜在生物标志物,鉴定该生物标志物的有效性,最终代替成为新的生物标志物。结合多种方法,如影像学及基因技术,对偏头痛临床亚型进行分类,提高精确度,并为精准医疗作出贡献。偏头疼领域的研究目前已到达需结合影像学发现及基因研究的地步。解决二者之间的连接问题可能提高治疗效果、早期介入使偏头疼不进入慢性化过程等。
方法整合
一些基因已证明与一些定义明确的偏头疼亚型相关(如偏瘫型偏头疼),同样存在MRI研究。但是这些研究并不能将存在割裂的研究结果联系在一起:MRI研究发现多脑区在疾病中受累,而GWAS研究发现变异基因与血管平滑肌相关。显而易见未来的研究方向应致力于将二者割裂的结果整合在一起,并明确其与偏头痛易感性、疾病亚型、药物治疗效果及并发症之间的关系。
医学及生物学研究倾向于大数据采集与分析,从生物系统模式理解疾病,如基因组学研究及脑影像学的行为过程研究。上述大数据研究方法已有许多有关人脑的研究,例如人类连接组计划。相较于基因研究,影像学大数据采集更加困难,因此,结合并分析基因数据及影像数据目前困难重重。尽管如此,脑影像学研究仍为一种可高度重复的方法,并且应用广泛,可与基因组学数据结合研究脑结构与功能;这些方法可用以研究偏头疼。随着结合二者的技术应用越来越广泛,这些技术可以用来发展疾病预测、疾病恢复及药物疗效的基因及影像学标志物。例如,决定树突棘密度的基因可能与偏头疼患者的大脑形态动态改变相关,并与药物治疗疗效相关。
结论及展望:
近年来针对偏头痛脑功能及基因相关性的研究结果让我们对偏头痛有了更多的认识。总的来说,神经影像学研究为我们提供偏头疼发作周期中脑功能、结构及化学改变的重要信息。这些研究的目的在于将临床所观察到的各种偏头疼亚型与影像学发现相关联,并发现新的神经影像标志物。fMRI及PET对于动态及静息态偏头疼患者大脑的研究为我们对于偏头疼发生及发展的理解提供显著的帮助,但多数相应研究仅为描述性研究。随着未来纳入更多样本的影像学研究发展,更多的标志物可能被发现。迄今,对于偏头痛GWAS研究的结果对我们今后精简研究设计提供帮助,并且为偏头痛血管功能提供新的焦点。更大规模的GWAS研究可能发现更多的偏头痛风险基因位点;未来更大的GWAS研究样本可能由大型商业或公立的数据库中。
新近的来自商业数据库的基因组学研究已发现更多新的风险基因位点。尽管研究成功发现多种风险基因位点,整合这些研究发现并非简单,表明偏头疼具有异质性特点。这些异质性在大样本数据研究中更加显著。
为了明确这些基因位点的致病性变异,未来基于特定偏头痛亚型的相关基因功能的研究需要进一步探索。结合基因及影像学数据研究将对进一步理解目前偏头疼研究结果提供帮助,并可能对未来进一步研究带来启发。对于特定基因结合影像学的研究可以用ASTN2基因多态性研究为例,ASTN2基因影响胶质介导神经元迁移过程,并可能与影像学发现的海马体积变化相关。相似的结合基因及影像学的研究潜力巨大。目前已有例如IHGC及ENIGMA等组织应用结合两种技术的方法研究偏头疼基因与MRI皮层下、颅内及海马体积等的相关性。
这些大型研究具显著的统计可信度,并可能确定全局的影响及特定SNPs对于偏头疼风险及脑功能的影响。“偏头疼组学”研究的发展将偏头疼研究带入到了影像学及基因组学结合的层面,并对偏头疼的复杂性及多样性的了解提供帮助。这种研究方法对于具有明确基因特征的偏头疼提供帮助最大(例如FHM:家族遗传性偏瘫性偏头疼),对于其他散发性偏头疼也可能发现新的标志物。成功将影像学及基因技术结合的研究将会加深我们对于偏头疼发病机制的理解并提高我们对偏头疼治疗的把握。并且可能帮助我们提供新的偏头痛缓解通路,发现新的药物靶点。一旦发现新的标志物,进一步的研究应当致力于发掘其所具有的诊断性及预测性价值。
如需原文请加微信:siyingyxf 或者19962074063获取,如对思影课程感兴趣也可加此微信号咨询。

微信扫码或者长按选择识别关注思影
非常感谢转发支持与推荐
欢迎浏览思影的数据处理课程以及数据处理业务介绍。(请直接点击下文文字即可浏览思影科技其他课程,欢迎报名与咨询):
第六届任务态fMRI专题班(重庆)
第十四届磁共振脑网络数据处理班(重庆)
第九届磁共振脑影像结构班(南京)
第十九届脑电数据处理中级班(南京)
招聘:脑影像数据处理工程师(重庆&南京)