

引言
创伤后应激障碍(PTSD)的主要特征是那些无法控制和令人痛苦的侵入性记忆的持续表达。在发生创伤事件之后,试图抑制或避免创伤记忆有时反而会增加侵入性记忆的表达。介入式记忆的成功治疗涉及克服这种避免和抑制,以及通过整合带回创伤式记忆的要素以促进其消失或更新。这些治疗方法符合当前的神经生物学模型,该模型将PTSD与学习障碍以及在恐惧回路中处理情境提示物的缺陷联系在一起。PTSD的理论暗示着通过抑制对维持入侵症状不利且至关重要的思维来避免创伤记忆的发生。
要求人们抑制由提示线索触发的记忆的意识而不诉诸该记忆,会破坏之后的有意识的回忆、无意识的表达或情绪反应。记忆抑制涉及由额顶网络参与的控制机制,抑制记忆提取会降低大范围网络的大脑活动。动机性遗忘的神经生物学模型假设,一旦提取线索触发了与意外入侵相关的干扰活动,对记忆意识的抑制控制就会自适应地抑制记忆加工。表现为右侧背外侧前额叶皮层(DLPFC),特别是前额中回(MFG)对负责记忆的大脑区域重新激活的影响。
这种自上而下的抑制作用增加,可以自适应地抵消和控制自主出现在意识中的入侵记忆。这些抑制控制机制的改变可能是导致PTSD个体不良反应的侵入性症状的基础。因此,在抑制尝试之后,PTSD中的侵入性记忆的持续可能是受损且无效的记忆控制系统引起的,进而削弱了分配其他必要的抑制技巧的能力。
本文作者检测了102名2015年经历巴黎恐怖袭击的个体(创伤暴露者)以及73名健康对照者的控制系统与记忆回路之间的功能连接(Fig. 1B)。根据DSM-5标准将102名创伤暴露者分为有PTSD症状组(PTSD,55名)和无TSD症状组(non-PTSD,47名)。在学习word-object对后,试图在think/non-think(TNT)阶段阻止object的记忆进入意识(non-think)。将抑制过程中回忆到该object的trial剔除,并在trial结束后报告提示物诱发的配对object感知,从而使研究者能够分离何时non-think trials触发入侵。
材料和方法
被试
88名未暴露者和120名暴露者参与该研究,其中暴露者招募自受灾者协会资助的跨学科的纵向研究“Programme 13-Novembre”。7个未暴露者被剔除:4个被试由于技术或行为问题没有获得侵入等级数据、2个被试的MRI数据有伪影、1个被试无法进行试验。18个暴露者被剔除:8个被试由于技术或行为问题没有获得侵入等级数据、3个被试在MRI中被中断、7个被试不满足纳入标准(6个被试符合重新出现症状的标准,但是不存在其他症状类别;1个被试实际上没有经历恐怖袭击)。
最终样本包含102名巴黎恐怖袭击的暴露者和73名未暴露的健康对照者,所有被试年龄在18-60岁、右利手、讲法语、体重指数< 35 Kg/m2。通过医生的临床检查以确保所有被试没有神经、医学、视觉、记忆或精神疾病。14名暴露者试验时在服用抗抑郁、抗焦虑或催眠药物(详细说明见S15)。将这些接受药物治疗的被试纳入分析能够反映一般的PTSD人群,并且还进行了协方差分析,以确保主要发现不依赖于这些被试。
暴露者由受过训练的心理学通过DSM-5结构化临床访谈(SCID)诊断,并受精神科医生监督。所有暴露者均符合DSM-5标准A,表明他们经历了创伤事件(详细暴露类型见table S1)。如果被试满足DSM-5定义的所有其他诊断标准,则将其标记为完整形式的PTSD(29名)。如果被试再次出现症状(标准B)并且持续超过一个月(标准F),并导致严重的困扰和功能障碍(标准G),则被诊断为部分形成的PTSD(26名)。尽管诊断为部分形式PTSD的被试未达到PTSD的全部临床症状,但在其中发现的侵入性症状导致严重困扰,可能造成了与完全PTSD相当的社交和功能障碍。
将完全形式PTSD和部分形式PTSD的暴露者分在一组进行统计分析,将其称为PTSD组的一个临床组。因此,该研究包含55名有PTSD的暴露者(PTSD+)、47名没有PTSD的暴露者(PTSD-)和77名未暴露者(Control)。
使用创伤后应激障碍检查表(PCL-5)评估PTSD症状的严重程度,此外使用状态特质焦虑量表(STAI)和贝克抑郁量表(BDI)分别评估焦虑和抑制程度。使用匹兹堡睡眠质量指数和失眠严重程度指数分别评估被试实验前一个月内的睡眠习惯以及是否失眠。
对四组被试正常睡眠时间与MRI采集前一晚的睡眠时间进行ANOVA分析,发现睡眠时间的主效应(F1,158 = 13.43, P < 0.001),而不存在组间交互作用(F3,158 = 0.02, P = 0.996),这说明所有被试MRI前一天晚上睡眠时间不正常,睡眠时间短。事后比较显示未暴露者比完全PTSD和(P = 0.03)非完全PTSD者有更长的睡眠时间(P = 0.013),但是在PTSD中不存在组间差异(P > 0.03),人口统计学和临床特征见table S1。
所有被试在2016.6.13 -2017.6.7之间完成研究,暴露者在巴黎袭击发生日期和纳入研究日期之间的延迟没有显着差异。该研究得到区域研究伦理委员会的批准。所有被试在参加之前均应遵守法国的道德准则,签署书面知情同意书。要求被试在实验前或实验期间不要服用精神兴奋剂、药物或酒精。
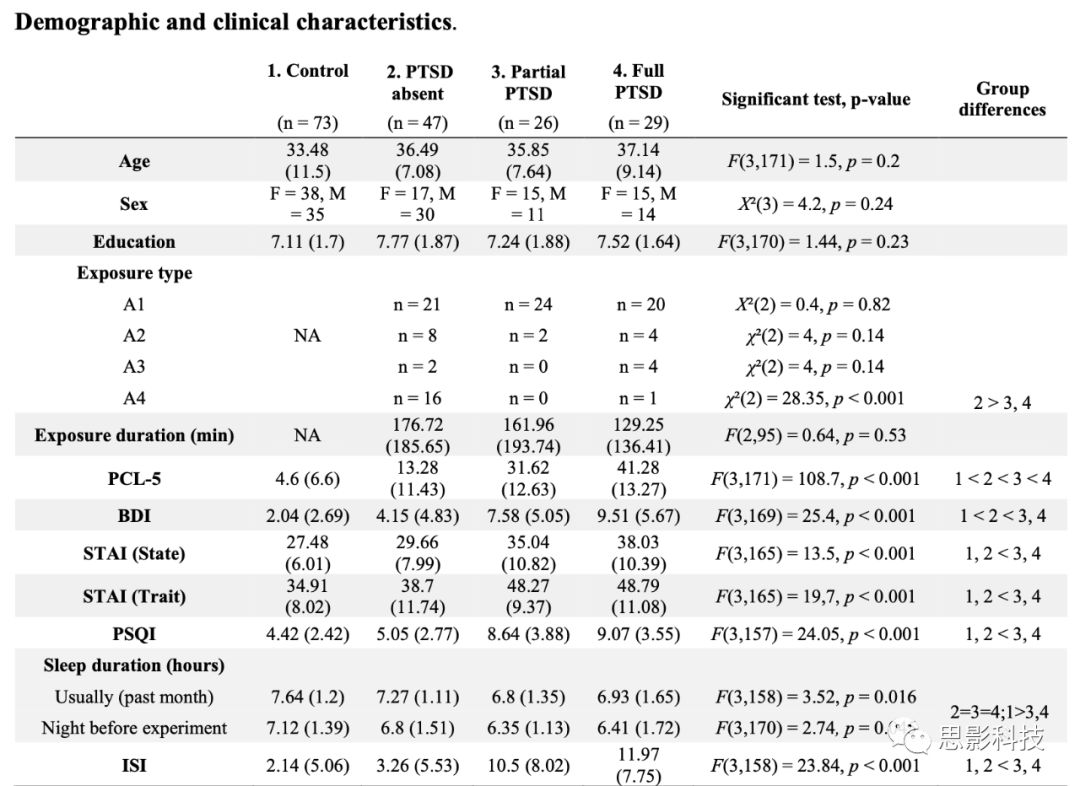
Table S1 人口统计学和临床特征统计指标
刺激是由72个单词对组成的三个系列列表,选自中性的抽象法语单词和标准刺激库(BOSS)。创建了三个系列,分为18对的四个列表,分别分配给四个条件(think,no-think,baseline和unprimed),外加8个用于练习的填充物。配对的列表在三个系列、四个条件和三组被试中平衡,并根据可能影响任务表现的不同属性进行了匹配。单词列表在平均命名延迟、字母数量和词频上匹配。对象列表对命名延迟、熟悉程度、视觉复杂程度、视点、名称和对象一致以及可操作性进行了匹配。使用与创伤经历完全脱节的中性材料使得能够研究一般的记忆控制机制,并避免创伤者的道德问题。
流程
在MRI数据采集之前,被试学习72个法语中性word-object对,每对呈现5 s。然后给定对的word在屏幕上呈现4 s,询问被试是否可以回忆并完全可视化与之配对的object(见图1)。如果被试确定可以回忆,接着在屏幕上呈现三个object(1个正确,2个错误),被试需要在4 s内选择与这个word相关联的object。每次识别测试后与word正确关联的object呈现2500ms,要求被试使用这种反馈来增加对知识的了解,直到准确率达到90%或者最多回忆6对。最后使用最终标准测试对所有记忆对进行了评估,但是未给出任何反馈,各组之间不存在差异(P > 0.18),即表明这种word-object关联学习在不同组中得到匹配。学习之后将72个word-object分为三个列表,分别分配给TNT(think/no-think task)任务的think、no-think、baseline条件。
在T1 images扫描过程中,在屏幕上再次呈现上述word-object对以强化学习。这种过度训练过程在于确保提示word会自动关联配对的object,从而使我们能够分离出在TNT阶段参与控制配对object侵入的大脑区域。
TNT任务分为4个session,每个session大约8分钟,其中think和no-think物体呈现两次,对应的提示word分别用绿色和红色呈现。在think trials中,被试需要尽可能详细地想象关联的object;在no-think trials中,被试需要阻止该object浮现在脑海中,并集中注意于提示word。每个trial结束之后被试通过按键反应报告关联的object是否在进入意识。使用遗传算法工具箱用于优化think和no-think对比的效率,添加20%的无 duration空事件以及时间伪随机呈现的注视屏。被试在TNT任务后进行知觉辨别任务以测试先前的抑制尝试是否会影响重复启动。
在每个trial中,每个item依次呈现,被试需要仔细观察item是否逐渐被打乱,并在能够看到并命名图片中的物体时尽快按键。通过使用傅立叶变换将图片分解为相位和幅度谱来实现加扰。图片在每个噪声级别上呈现150 ms,产生的总刺激持续时间为3.15 s。两次trial之间的平均刺激间隔为2.4秒,并且还增加了20%的无效事件。最后采集被试的静息态数据,提示被试闭眼休息,并保持静止和清醒。最后,使用调查问卷询问被试在TNT阶段使用的策略。结果显示,被试仍然专注于屏幕上显示的word,并通过直接压制关联的object来控制不需要的记忆。与其他抑制策略相比,被试对控制no-think项目的认知要少得多(z > 140,P < 0.001)。此外,Kruskal-Wallis检验说明使用任何一种策略的组之间没有任何差异[H(2)< 2.73,P > 0.26]。
MRI数据采集
使用3T Philips磁共振扫描仪收集数据,首先采集T1 images:TR = 20 ms, TE = 4.6 ms, flip angle = 10°, multi-band采集,SENSE factor = 2, 180层, voxels = 1 mm*1 mm *1 mm, FoV = 256 mm*256 mm *180 mm, matrix = 256 mm*130 mm*180mm;然后采集TNT任务和静息态数据:TR = 2050 ms, TE = 30 ms, flip angle = 78°, 32层, 层厚3mm, 层间距0.75mm, matrix = 64 mm*64 mm*32mm, FoV = 192 mm*192 mm *119 mm。
fMRI预处理
使用SPM12软件进行头动校正、时间层校正、与颅骨剥离后的T1配准。将T1分割为灰质、白质和脑脊液,并非线性标准化到MNI空间。形变场中的每一个点是MNI标准空间与原始空间以毫米为单位的映射,使用该映射将MNI标准空间的ROI坐标投影到原始空间。以此后续的所有分析均在未进行空间平滑的原始空间中进行。
Think/no-think单变量分析
对经预处理的数据进行高通滤波及协变量去除。填充事件以及think条件下未按键、有按下按钮但错误回忆的trial被去除。通过将每个感兴趣条件的first-level激活图输入SPM进行基于体素的分析。对于contrast:no-think - think在全脑范围内进行统计检验,阈值设定为PFWE < 0.05;对于contrast:intrusion(侵入条件) – no-intrusion(非侵入条件)的组内及组间比较在合适的感兴趣volume内进行,阈值为Puncorr < 0.005。使用回归模型分别在各组内研究大脑活动与侵入频率之间的关系,Puncorr < 0.005。
Regions of interest (ROIs)
本文集中于TNT研究中报道的前额叶和记忆系统,分别对应于up-regulate(正向调节)和down-regulate(负向调节)对不想要记忆的抑制。基于Brainnetome atlas选择ROIs,该模板将全脑划分为210个皮层区域和36个皮层下区域。考虑到前额叶控制网络在记忆抑制中存在较强的右半球偏侧化,本文选择记忆提取抑制中一致激活的脑区:右侧额上回SFG、右侧额中回MFG、右侧额下回IFG、右侧前扣带回CG。从记忆网络中,本文选择记忆抑制任务中一致被抑制的双侧区域:海马旁回、旁海马旁回、梭状回、腹侧楔前叶。基于前额叶控制网络的20个ROI和记忆网络的18个ROI(见table S8),构建个体的功能连接矩阵。为了进行组间比较,本文选择右侧MFG的前部区域(见补充材料table S8,太长,这里不贴出了,大家可以在补充材料原文中查看)。
功能连接分析
本文使用广义形式的PPI(gPPI)研究前额叶控制网络和记忆网络区域内(见table S8)与任务相关的功能连接,关注抑制侵入刺激与非侵入刺激之间的对比。本文的设计通过较短的刺激间隔和主要条件之间的缓慢变化来优化条件之间的信号变化检测。为了减少被试在任务态下,在没有刺激的情况下采集的数据的时间很短。为了解决静息态基线可能被任务的认知过程污染,本文使用盲反卷积方法检测在TNT任务之后静息态信号与事件相关的自发变化。对TNT和静息态的每个ROI的BOLD时间序列进行反卷积估计神经活动。将估计的神经活动与编码TNT或静息态的Boxcar函数相乘来构建PPI回归,然后对PPI回归去趋势及归一化,以方便在TNT和静息态之间进行比较。
有效连接分析
本文使用DCM和贝叶斯模型平均法评估在抑制侵入性记忆过程中,由于控制要求增加而导致的右侧MFG和记忆系统之间的连接调节是否是由自上而下的过程介导的。通常认为记忆抑制来源于右侧MFG的前部,因此本文关注右侧MFG对同侧半球内记忆区域的影响。由于DCM分析需要控制一定数量的节点,本文关注内侧颞叶MTL(包括海马旁回、旁海马旁回)和楔前叶。
由于在PPI分析过程中caudal海马旁回没有明显的调控作用,因此在DCM分析中排除此区域。本文构建了七个DCM模型及一个空模型(见图5A),将该空模型与其他调制模型相比较以确保抑制引起连接的某种调制。在估计8个模型后,首先通过贝叶斯模型选择(BMS)比较调制模型与空模型,结果显示BMS压倒性地偏向于调制模型,对未暴露组、non-PTSD组和PTSD组,超越概率EP = [100%, 100%, 100%],预期后验概率EPP = [91%, 88%, 78%]。
统计分析
对于记忆抑制诱导的功能连接改变,使用单尾配对样本t检验进行组内比较、单尾双样本t检验进行组间比较,阈值为PFDR = 0.05并假设存在条件组间的正趋势。此外,作者使用马尔可夫链蒙特卡洛方法(MCMC)的贝叶斯方法,估计贝叶斯因子(BF)以用于可视化来表示每个组内的抑制作用的可能性。基于此假设,我们将实际等效区域(ROPE)定义为Cohe’s d效应大于-0.1的区域。MCMC方法生成了50000个可靠的参数组合,这些组合代表了后验分布;然后估计ROPE中后验比例与ROPE中先验比例的比率。对于ROI分析,使用非参数置换检验,对被试集进行5000次迭代并计算95%置信区间来检验组内差异。使用ANOVA模型,在控制年龄、性别、教育程度、药物治疗、创伤暴露的类型、持续时间后进行组间比较。对于DCM,BMA提供每个组耦合参数后验分布的均值和标准差。与DCM贝叶斯框架一致,估算了组内和组间差异的后验概率和95%置信区间。
行为学表现
如图1C所示,在所有三个组中,通过重复抑制的方法,被试对侵入的控制有所提升。对被试报告no-think trials进行ANOVA分析显示,侵入的比例随着重复抑制显著降低[F7,1204 = 30.3, P < 0.001],在三个组中均有减少[F14,1204 = 0.46, P = 0.95],而且侵入的总比例不存在组间差异[F2,172 = 2.1, P = 0.125]。
相比于基线条件,未暴露者 [t72 = 1.96, P = 0.027]以及non-PTSD [t46 = 1.73, P = 0.045]对no-think object的识别更慢(见table S2),即priming effect降低。记忆抑制之后,priming effect在PTSD不显著[t54 = −0.84, P = 0.4],而与PTSD相比,该效应在未暴露者[t100 = 1.85, P = 0.033]和non-PTSD[t126 = 1.95, P = 0.027]中显著更大。本文的训练程序已经在不同组间得到匹配,因此上述结果说明,PTSD个体在重复抑制训练后,对由词汇诱发的外显记忆的遗忘受到了调节的明显影响,本研究的发现将这种缺陷扩展到了感知的内隐记忆。
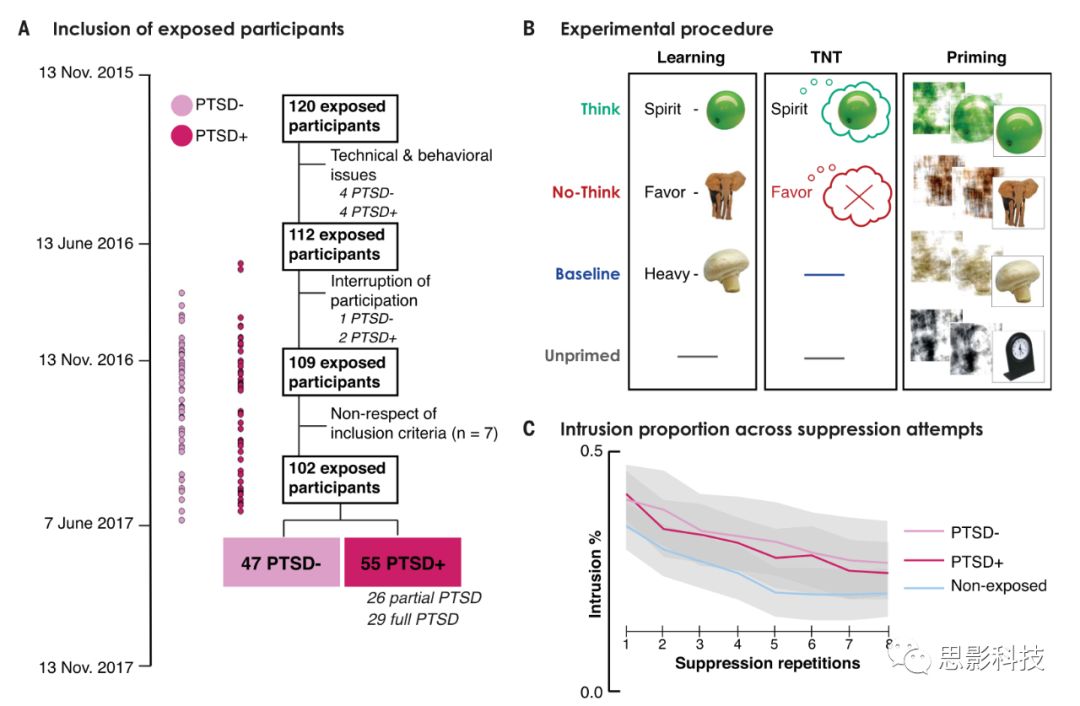
图1 实验设计
(A)经历2015年巴黎恐怖袭击的被试的时间和程序。non-PTSD组和PTSD组有相似的暴露程度。
(B)在学习word-object对后,收集被试完成TNT任务并接受磁共振扫描。对于think trials(绿色所示),被试需要回忆与word相匹配的object。而对于non-think trials,被试需要阻止object进入意识。non-think trial结束后,被试需要报告侵入性记忆是否存在,这些侵入性记忆进一步触发了反应抑制过程。
(C)入侵比例,即non-think trials中相关记忆进入意识的比例。
脑活动
首先在全脑范围内比较了non-think trial与think trial大脑活动,在记忆提取的抑制过程中,右侧额顶控制网络FPCN的大脑活动增强,视觉区域和内侧颞叶MTL区域的大脑活动减弱,未发现non-PTSD与PTSD的组间差异。
但是当研究范围限制到FPCN时,与未暴露组相比,在PTSD组发现交互作用,且该交互作用来自于未暴露组右侧额上回SFG(x, y, z = 16, 36, 56; Z = 4.34, PFWE-FPCN = 0.002)更高的大脑活动。目前不清楚上述结果来自于PTSD或者是创伤暴露。这种交互作用也可能反映创伤个体在记忆控制过程和某种习惯形式的参与。此外,创伤相似区域的皮质厚度增加,可能与经验诱发的可塑性和习惯化有关。进一步探究了个体抑制入侵记忆的能力是否依赖于FPCN的参与。在未暴露组和non-PTSD组均发现FPCN增强的活动与减少的入侵频率有关(fig. S2)。虽然双侧海马旁回大脑活动降低,但是不受入侵相关的控制要求调制。在non-PTSD组与PTSD组,视觉系统外侧及后侧区域降低的大脑活动与入侵抑制有关。
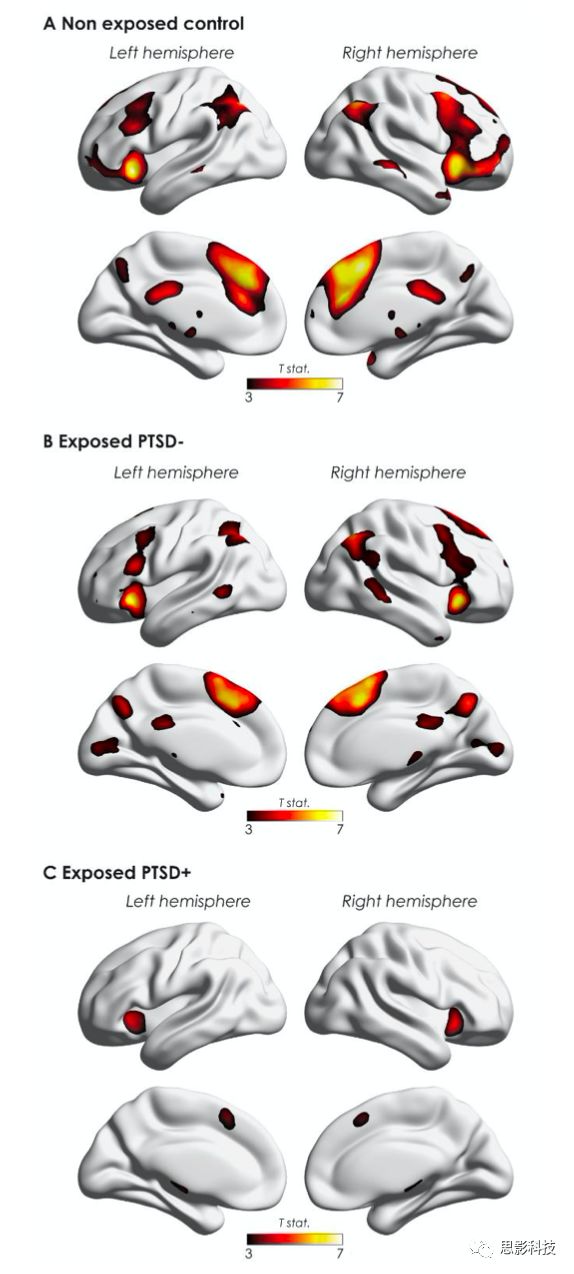
Figure S2 FPCN增强的激活和侵入频率有关
功能连接
本文接下来研究了抑制控制网络与记忆区域之间的功能连接模式(Fig. 2A、table S8),其中抑制控制网络包括右侧DLPFC及前扣带皮层。对未暴露组与non-PTSD组,与非侵入性记忆相比,阻止入侵性记忆进入意识的尝试与大范围功能连接的显着降低有关(Fig. 2B),来自于侵入过程中额叶网络与旁海马旁回、海马旁、梭状回和楔前叶之前功能连接的降低,在PTSD组未观察到功能连接的降低(Fig. 2B)。我们进一步关注右侧前MFG与记忆区域之间功能连接的改变(Fig. 2A),结果显示与PTSD组相比,non-PTSD组功能连接降低的区域在右侧海马旁回、双侧旁海马旁回、右侧梭状回、双侧楔前叶。这些差异与创伤暴露的类型、持续时间、年龄、性别、教育程度、药物治疗无关。对于未暴露组,与非入侵条件相比,入侵条件下功能连接显著降低的区域在双侧楔前叶。入侵条件-非入侵条件后,与PTSD组相比,未暴露组左侧旁海马旁回、双侧梭状回、左侧楔前叶功能连接显著降低,右侧海马旁回有趋势,而只有左侧楔前叶能够通过FDR校正。在控制年龄、性别、教育程度、药物治疗后,左侧楔前叶仍然显著。
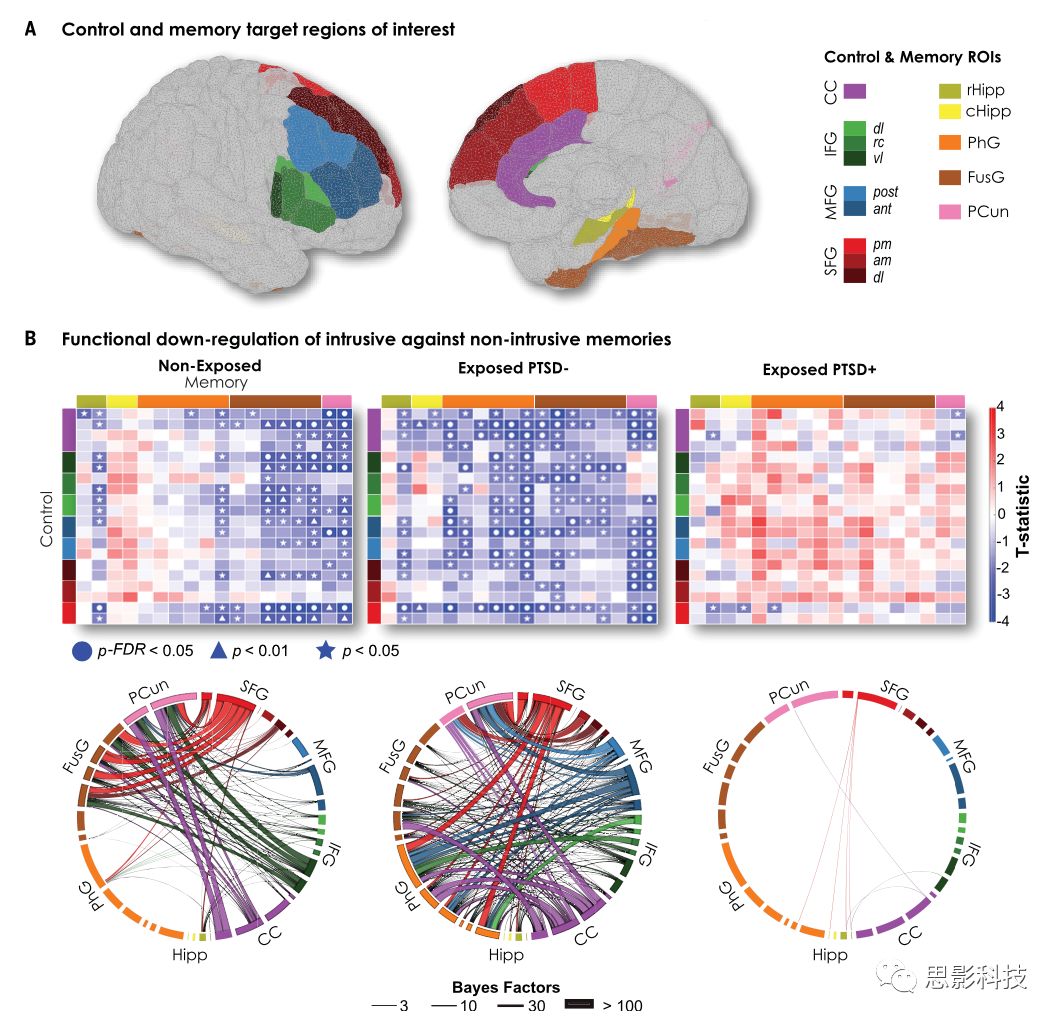
图2 在抑制侵入记忆的过程中,控制区域和记忆区域之间的功能连接降低
(A)前额叶控制区域和记忆区域之间记忆诱导的功能连接。
(B)入侵记忆与非入侵记忆对比后,未暴露组和non-PTSD组大范围功能连接降低。
矩阵表示每一组控制系统和记忆系统ROI之间的功能连接变化。○、△、分别代表阈值在PFDR < 0.05、P < 0.01、P < 0.05下的显著变化。在圆形连接图中,边的颜色由前额叶控制区域ROI定义,边的大小反映了与非入侵记忆相比,在抑制入侵记忆的过程中与功能连接显著降低相关的连接的贝叶斯因子。

图3 记忆抑制过程中前MFG与记忆系统之间的连接调制在抑制入侵记忆与非入侵记忆过程中,右侧前MFG(seed)与记忆系统目标区域(上图:左半球,下图:右半球)之间的功能连接差异。黑色和白色分别表示阈值为PFDR < 0.05和P < 0.05。
任务态功能连接与静息态功能连接
进一步将每种情况下与任务相关的功能连接与静息态功能连接(TNT后采集)比较。以静息态功能连接为基线,作者在未暴露组与non-PTSD组发现,入侵条件下右侧DLPFC区域与记忆区域功能连接降低(fig. S5)。在PTSD组发现相似的功能连接降低,但是主要在非入侵条件。此外,未暴露组在非入侵条件也观察到功能连接降低,说明在在在非入侵条件和入侵条件下未暴露组均能抑制记忆活动。对于未暴露组与non-PTSD组,入侵条件下记忆区域(包括海马旁回)与右侧MFG之间的功能连接降低(Fig. 4)。
对于未暴露组,双侧梭状回、右侧海马旁回也观察到功能连接降低;对于PTSD组,左侧旁海马旁回、右侧梭状回观察到功能连接降低,虽然这些区域未通过多重比较校验。未暴露组由记忆抑制所诱发的抑制控制网络与记忆区域之间的功能连接降低反映了一个主动的过程,即当侵入性记忆进入意识并需要被试清除时,这些功能连接增加。这些结果与当前的动机性遗忘的神经生物学模型非常吻合,该模型认为一旦回忆提示意外触发了干扰性侵入活动,对记忆的抑制控制会自适应地增加以抑制记忆加工。
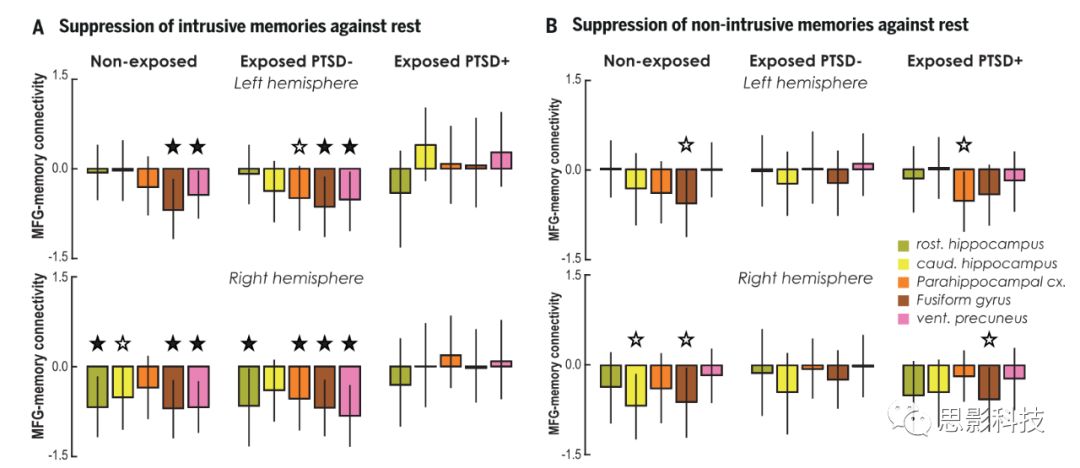
图4 抑制诱发的功能连接与静息态功能连接以右侧前MFG为seed、记忆区域为target,抑制入侵记忆
(A)与非入侵记忆(B)诱发的功能连接与基线(静息态功能连接)的差异。黑色和白色分别表示阈值为PFDR < 0.05和P < 0.05。
自上而下(top-down)与自下而上(bottom-up)连接
使用动态因果模型DCM分别分析了抑制入侵性记忆过程中自上而下(top-down)与自下而上(bottom-up)的影响。本文DCM的节点限制于右侧前MFG、海马旁回、旁海马旁回和楔前叶(Fig. 5A)。使用贝叶斯模型平均(BMA)根据MFG和记忆区域之间所有七种可能的调制组合的后验模型衡量耦合参数的改变。对于non-PTSD与未暴露组,参与侵入性记忆抑制的海马旁回受右侧前MFG自上而下的调控。在PTSD组未发现这样的调控,来自于抑制非侵入性记忆过程中,右侧MFG-海马旁回之间功能耦合的下降。在PTSD组与non-PTSD组、PTSD组与未暴露组,均观察到右侧MFG-海马旁回之间自上而下耦合参数的组间差异。与PTSD组与未暴露组相比,non-PTSD组显示出楔前叶自下而上的调控。与其他组相比,non-PTSD组观察到的自上而下的连接的降低与创伤暴露的类型、持续时间、年龄、性别、教育程度、药物治疗无关。
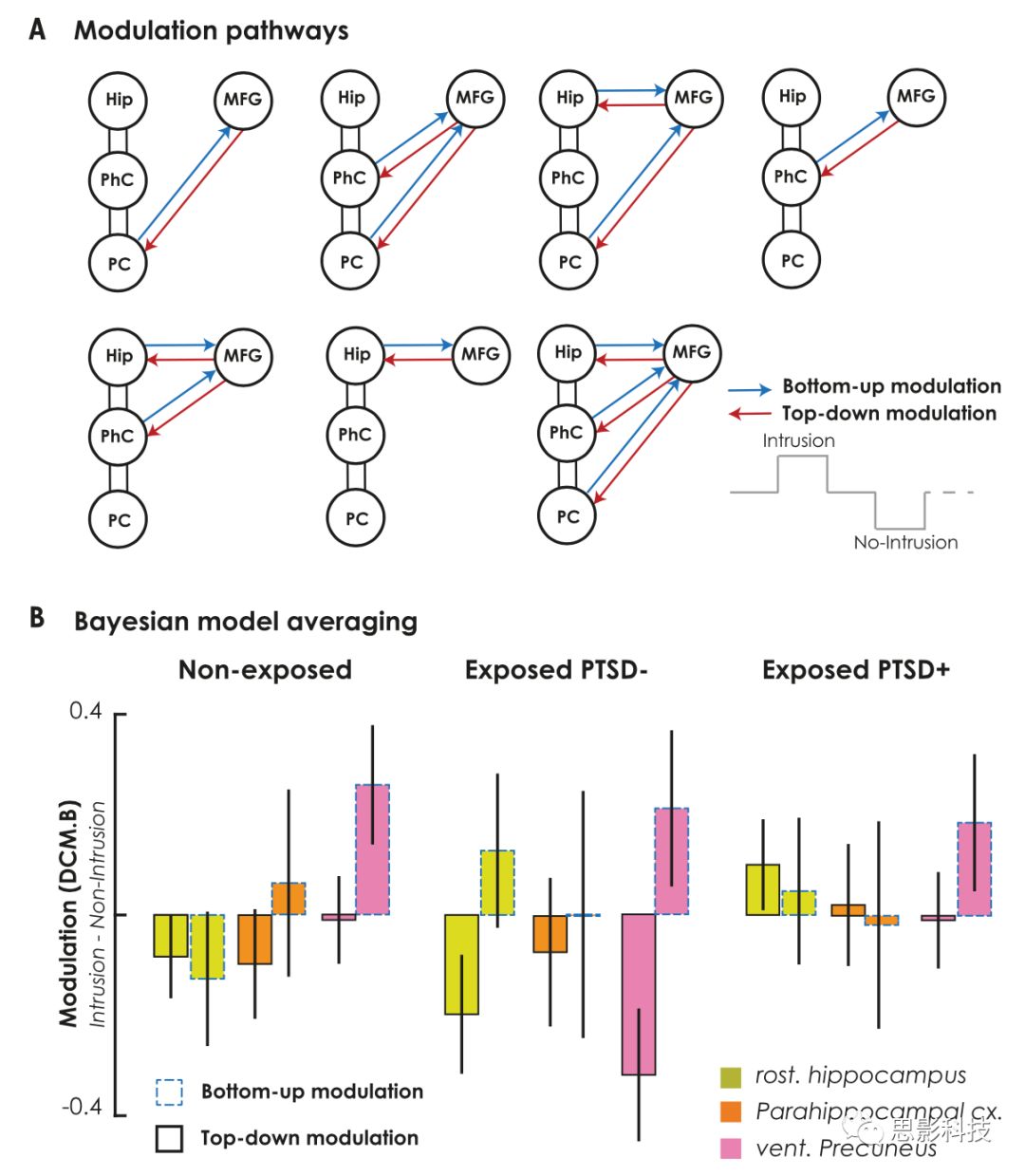
图5 DCM模型与耦合参数
(A)抑制尝试过程中右侧前MFG与记忆区域之间自上而下top-down、自下而上bottom-up的影响。作用在这些连接上的调制输入反映了入侵记忆与非入侵记忆之间耦合的差异。记忆目标区域包括海马旁回Hip、旁海马旁回PhC和楔前叶PC。
(B)在自上而下和自下而上的调制参数的模型空间中的贝叶斯模型平均。
PTSD患者对侵入性记忆的抑制控制普遍存在缺陷
当前的PTSD模型将侵入性记忆的持续性与创伤后遗忘或更新创伤记忆痕迹的失败联系在一起,同时异常及夸大的处理恐惧回路中创伤的情景提示。这些异常与海马旁回-杏仁核的功能障碍及其与内侧前额叶皮层的相互作用有关。本文结果表明,PTSD的特征还在于与侵入性记忆有关的自上而下地抑制瞬时意识的缺陷,这种缺陷可能是创伤性记忆持续存在的中心因素,削弱了分配维持健康记忆所需资源的能力。
在non-PTSD个体中在调控侵入性记忆的过程中(相比于非侵入性记忆),涉及抑制控制的前额叶区域与记忆区域(海马旁回、楔前叶)之间的功能连接降低。在与静息态数据的比较中也发现上述连接的降低,表明由抑制侵入性记忆诱发的功能连接的改变依赖于主动调控。有效连接分析显示,在non-PTSD个体中自上而下的过程介导了这些调节,而且与PTSD组相比这种影响在non-PTSD组更加明显。上述发现与抑制信号的存在相一致,该信号破坏了记忆系统中不必要的记忆痕迹跟踪的重新激活,这种抑制控制在有忍耐力的个体中得以保留,但在患有PTSD的个体中被破坏。虽然本文创建的侵入性记忆与PTSD中出现的令人痛苦的、破碎的创伤性侵入完全不同,但是对PTSD症状来说很重要的共同特征均存在,并且可以使用TNT范式进行建模和分离。因此,本文的发现凸显了PTSD个体前DLPFC区域抑制功能的中心性和普遍性破坏,甚至破坏了对中性的、人为创造的在学习中强度相等且完全不相关的无创伤事件的控制和抑制。
通常认为抑制记忆是不明智的,因为不想要的残余记忆会适得其反。而本次研究表明控制系统受损导致不适应的和失败的抑制尝试。此类破坏可能会阻止改变海马体的记忆稳定性的适应性遗忘过程,因此可能会阻止外伤性痕迹的损伤。此外,控制能力的改变会进一步加剧对创伤提示的避免。与记忆抑制不同(memory suppression),避免提示(avoidance of reminders)通过抑制控制、消灭或更新来阻止创伤表征的调节。抑制控制过程的中断可能会加剧记忆抑制和避免策略之间的不平衡,反映了虽然它们的目的都是使创伤记忆失去意识,但是对心理健康却产生相反的影响。
总结:
研究结果发现,在健康的个体(PTSD和未暴露的)中,试图阻止侵入性记忆的意外出现进入意识状态与控制系统和记忆系统之间的功能耦合显著降低有关,而在另一种情况下,任务cue并没有触发这种入侵。另外,对神经信息流动方向性的分析表明,健康个体对侵入性记忆的抑制源于右前区DLPFC的调节,后者调节了记忆过程的反应,以减少其反应。值得注意的是,这一调节针对的是两个与创伤记忆重新体验相关的关键区域:海马体和楔前叶。本文观察到创伤后应激障碍(PTSD)中控制不需要的记忆时其重新激活的调节信号的普遍中断。这种破坏可能是创伤性记忆持续存在的一个核心因素,削弱了部署维持健康记忆所需的应对资源的能力。这种缺陷可能解释了PTSD中常见的适应不良和失败的抑制尝试。本次研究表明,通常用于驱逐和抑制不想要的记忆的侵入性表达的一般心理活动可能有助于创伤事件后的积极适应,为新的治疗方法铺平道路。
原文:Resilience after
trauma:The role of memory suppression

微信扫码或者长按选择识别关注思影
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
更新通知:第十届脑影像机器学习班(已确定)