

ASD(孤独症谱系障碍)的研究目的之一就是确定早期生物标志,以指导生理病理诊断。EEG捕捉到的脑电振荡被认为是ASD生理病理学的核心。来自哈佛医学院的Laurel J. Gabard-Durnam等人在NATURE COMMUNICATIONS杂志发文,研究者以3-36月大的ASD高/低风险婴儿为被试,测量纵向EEG power,来探讨EEG power如何以及何时能够区分被试3岁时患ASD的风险以及是否患有ASD。第一年、第二年和前3年的EEG数据被放进数据驱动模型中来区分ASD。出生后第一年的动态功率能最有效地区分不同组别的婴儿。delta和gamma频段的功率轨迹能区分ASD婴儿和正常婴儿。此外,随着时间的推移也出现了一种发展趋势,高频段更易区分不同ASD症状。
早期的识别和干预对于改善ASD症状至关重要,不过ASD的明显行为症状一般在出生后第二年才开始显现,因此需要更敏感的指标来更早地识别ASD。研究表明,大脑振荡节律的早期中断是ASD的核心神经特征,而这些节律可以通过EEG捕捉。ASD高风险组和ASD患病组在不同脑电诱发振荡存在很多差异,特别是ASD高风险组的额叶EEG差异在婴儿早期就已出现。患ASD风险较高的婴儿在3个月大时就表现出额叶α和β频段的功率较低,6个月大时额叶δ到γ频段的功率较低,2岁时δ、α、β、γ等频段均存在差异,可见功率变化与年龄息息相关。与其他皮质区域相比,额叶脑电各频段功率的变化与ASD行为症状密切相关,如感觉低反应性、认知缺陷、语言以及受限和重复行为的程度。然而,早期EEG差异能否区分随后的ASD表现仍有待确定。
纵向研究发现出生后一年,婴儿全频段的EEG power出现了明显的改变,EEG power区分ASD症状的能力在这一阶段也会变化。此外,EEG power随发育产生的变化可能可以作为诊断措施,但迄今为止还没有研究表明是否可以利用脑电发展轨迹来区分ASD。因此需要进行纵向比较,以确定什么时候EEG power区分ASD最有用,以及EEG power生物标志在早期神经发育过程中是如何变化的。
本研究旨在探讨如何利用EEG power来判别出生后的3年ASD的患病风险和是否患有ASD。高风险人群具有较高的ASD发病率,如有兄弟姐妹患有ASD,这些有助于对ASD早期病理生理进行前瞻性检测。研究者分别多次采集ASD高风险婴儿和ASD低风险婴儿的静息态EEG,得到跨时间尺度的EEG数据。鉴于前人研究发现额叶各频段功率存在显著差异,因此重点分析额叶区域。重点分析三个主要的发育阶段:第一年,行为症状出现的前一年;第二年,出现新的行为症状;出生后前3年,包括ASD确认的年龄(3–12 months, 12–24 months, 3–36 months)。分析时,首先评估不同发育阶段EEG power鉴别的表现,以确定哪个发育阶段能最佳区分ASD的患病风险和是否患有ASD。此外进行补充分析,比较额叶皮层EEG与全脑EEG和顶叶EEG结果,以检验空间特异性。最后,哪些脑电频段及其发育轨迹图可作为潜在的生理诊断指标,以及这些频段在三个发育阶段是否发生改变。研究的最终目的在于通过测量和分析不同发育阶段的EEG power 来提供最鲁棒的鉴别ASD患病风险和是否患有ASD的指标。
方法
被试
本研究中,如果婴儿至少有一个被确诊为ASD的哥哥或姐姐,并且不能归因于已知的遗传病,那么就分到ASD高风险组(high-risk for autism,HRA)。如果婴儿有哥哥或姐姐,并且没有一级或二级家庭成员患有自闭症,那么就分到ASD低风险组(low-risk for autism controls,LRC)。结合自闭症诊断观察表(Autism Diagnostic Observation Schedule,ADOS)和临床诊断确定所有婴儿的ASD相关症状,36个月龄时确定了三组婴儿:ASD患病组,未患ASD的HRA组,未患ASD的LRC组。三组被试分布见表5。
ASD组的男性显著多于HRA(高风险)组(PearsonX2(1,102)=4.64,p=0.031)。LRC(低风险)组的父母教育水平显著高于ASD组(PearsonX2(2,87)=12.65,P=0.002)和HRA治疗组(PearsonX2(2,127)=6.62,p=0.036)。相应地,性别和父母教育水平作为潜在协变量纳入模型选择以确保EEG结果的差异与人口统计学变量无关。补充分析在没有性别和父母受教育程度作为潜在协变量的情况下进行,其结果与报告结果高度一致。
分别在婴儿3、6、9、12、18、24、36个月大时采集EEG数据。并非所有婴儿在所有时段都能采集到足够的脑电数据,可用数据的分布详见表5。
EEG数据采集
利用64导/128导Geodesic Sensor Nets采集静息态EEG数据,时间2到5分钟。高通滤波0.1Hz, 在线参考电极Cz, 阻抗保持在100KΩ下。婴儿在照料者的陪同下完成,采集环境符合标准研究条件。研究中的电极帽有64导和128导两种。
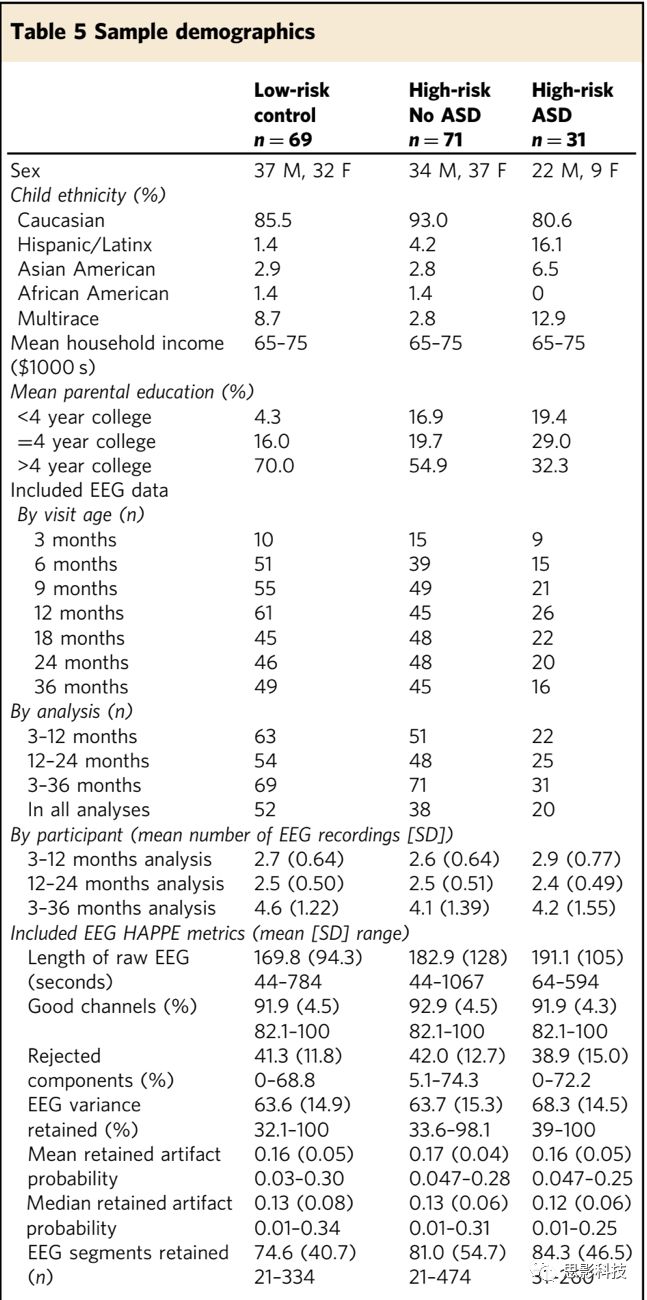
电极分布:
64-channel net—2, 3, 6, 8, 9, 11, 12, 13, 15, 16,17, 21, 24,25, 27, 28, 34, 37, 40, 46, 49, 50, 52, 53, 54, 57, 58, 61, 62;
128-channel net—3, 4, 9,11, 13, 19, 20, 22, 23, 24, 27, 28, 33, 36, 40, 41, 45,46, 47, 52, 58, 62, 70, 75, 83, 92,96, 98, 102, 103, 104, 108, 109, 112, 117, 118,122, 123, 124(详见补充图1)。
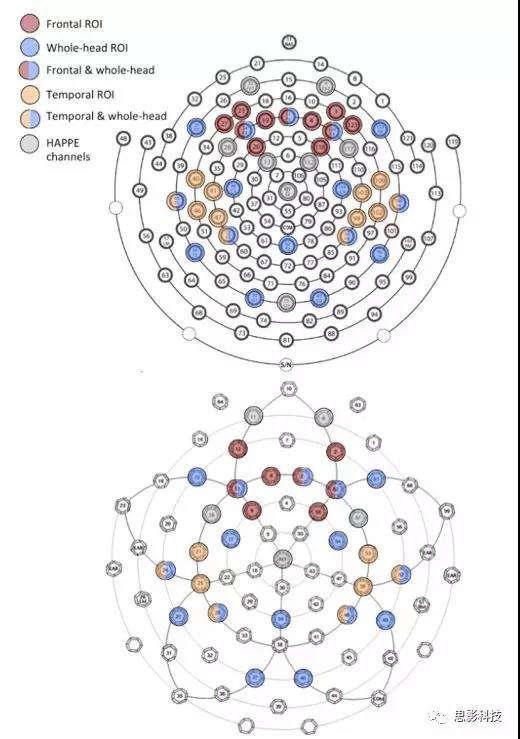
补充图1 电极分布
EEG数据处理
1、从NetStation中导出数据,并转换为matlab格式(R2017a)。
2、数据预处理、伪迹去除和数据质量评估在Harvard Automated Processing Pipeline for EEG (HAPPE)实现。批处理在batch electroencephalography automated processing platform (BEAPP) software实现。
3、进行1Hz高通滤波和100Hz低通滤波,去除60Hz的工频干扰。采样率从500Hz降到250Hz。利用EEGLAB的ICA功能去除眼动、肌肉活动等伪迹。对伪迹过大的通道进行球形插值。全脑平均参考,去线性漂移。分段2秒一段,剔除伪迹过大的分段。
EEG去除标准
标准的HAPPE处理会根据数据质量推荐合适的阈值,然后半自动化的剔除伪迹。总共40个分段,如果剔除伪迹后剩余的分段小于20,或者EEG的功率超过均值的三个标准差,则将该数据剔除。
EEG功率分解
对感兴趣通道的每个分段进行快速傅里叶变换。对位于额叶的通道进行初步分析(64-channel net—2, 3, 8, 9, 12, 13, 58, 62;128-channelnet—3, 4, 11, 19, 20, 23, 24, 27, 118, 123, 124)。分析频段如下:delta, 2–3.99 Hz; theta, 4–5.99 Hz;low alpha,6–8.99 Hz; high alpha, 9–12.99 Hz; beta, 13–29.99 Hz; gamma, 30–50Hz。
对每个频段内的所有频率点求和得到总功率,总功率与该频段功率密度曲线下的面积相等(图2)。对每个被试每个通道每个分段数据求平均并进行log 10变换。每个频段的总功率为所有感兴趣通道功率的均值(图3)。
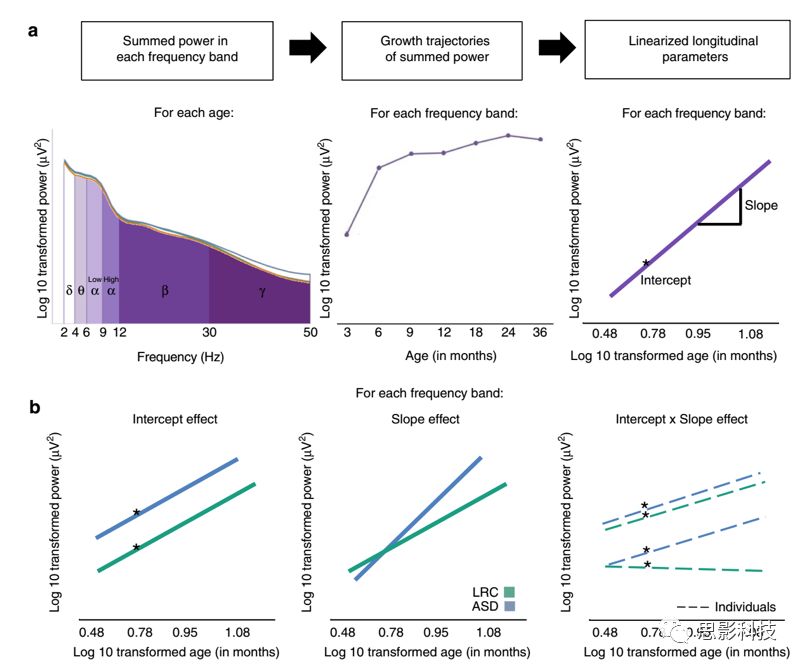
图2 分析示意图。该图阐明了如何生成和分析纵向EEG参数。
a (对于每个被试):计算每个年龄段每个感兴趣频段(delta, theta, low alpha, high alpha, beta, gamma)的总功率,其大小等同于EEG power的曲线下面积。绘制每个频段的EEG power随年龄变化的发育轨迹(本图中以beta在3-36月间的发育轨迹为例)。对每个频段总功率的发育轨迹按照年龄进行log10变换,这样就能求得随年龄变化的斜率和截距,用于后续的组分析。
b(组分析):在数据驱动模型中测试三组效应以区分不同的组别。本图中以low-risk control (LRC, in green color) vs.Autism(ASD, in blue color)为例,左侧图为截距的主效应,中间图为斜率的主效应,右侧图为斜率和截距的交互效应。
纵向EEG power(被试内)
利用SAS软件进行被试内分析,本研究用两个参数总结了每个频段EEG总功率的发展轨迹:EEG总功率截距和跨年龄的EEG总功率斜率(发育斜率,见图2)。为了获得这两个参数,研究者对每个婴儿的数据进行最小二乘回归(least squares (OLS) regressions),建立每个频段总功率随年龄变化的模型。将婴儿年龄、每个频段的EEG总功率进行log10变换,并建立模型观察二者的线性关系。这样只需要两次记录的脑电数据,就可以计算出这条线性回归线,并为组间分析提供数据。使用3个年龄段(3-12\12-24\3-36)的所有婴儿数据进行个体OLS(最小二乘)回归分析。12月份的数据同时纳入3-12和12-24,有助于样本量最大化,并能更完整的覆盖出生后第一年和第二年。
对于每个发展窗口,利用OLS回归方程生成每个数据在每个频段的总功率截距和线性发展斜率值。针对3-12月龄和3-36月龄的总功率截距分析,利用拟合回归方程计算每个婴儿6个月龄时的总功率。
数据驱动模型建构
感兴趣的预测因子包括EEG总功率的截距、EEG总功率的发育斜率、每个频段内截距与斜率的交互模型参数(一共18个总参数,详见图2)。鉴于儿童性别和父母教育程度不同,这两个变量也被输入作为潜在的预测因素。模型选择采用混合逐步选择法(即数据剖分法),其方法是最小化AIC准则。利用AIC准则识别最精确的、推广性最强的可区分不同组别模型。分别构建了三组比较模型:ASD与HRA组,ASD与LRC组,以及HRA与LRC组。分别用(A)3~12个月的EEG数据,(B)12~24个月的EEG数据和3~36个月的EEG数据构建二元Logistic回归模型。模型选择完成后,进行事后假设检验(多元Logistic回归中参数估计的Student’s t-tests),以确定每个模型中区分结果的相对鲁棒的参数。
模型比较的标准
为了比较随着年龄发展不同组别AIC最小逻辑回归模型的表现,使用被试内ROC曲线。首先,为了比较逻辑回归模型区分不同组别的准确性,计算每个模型95%置信区间的ROC曲线下面积(AUC),AUC指标在不同条件下是鲁棒的。使用最大化约登指数(灵敏度和特异度之和减去1。指数范围介于0-1之间。表示筛检方法发现真正病人与非病人的总能力)的阈值来评估模型的敏感性和特异性并进行约束。为了评估不同模型之间的真阳性和真阴性的分化率,研究者分别计算了对应于约登指数的阈值PPV(阳性预测值)和NPV(阴性预测值)。这些预测值解释了样本中虚假的患病率。
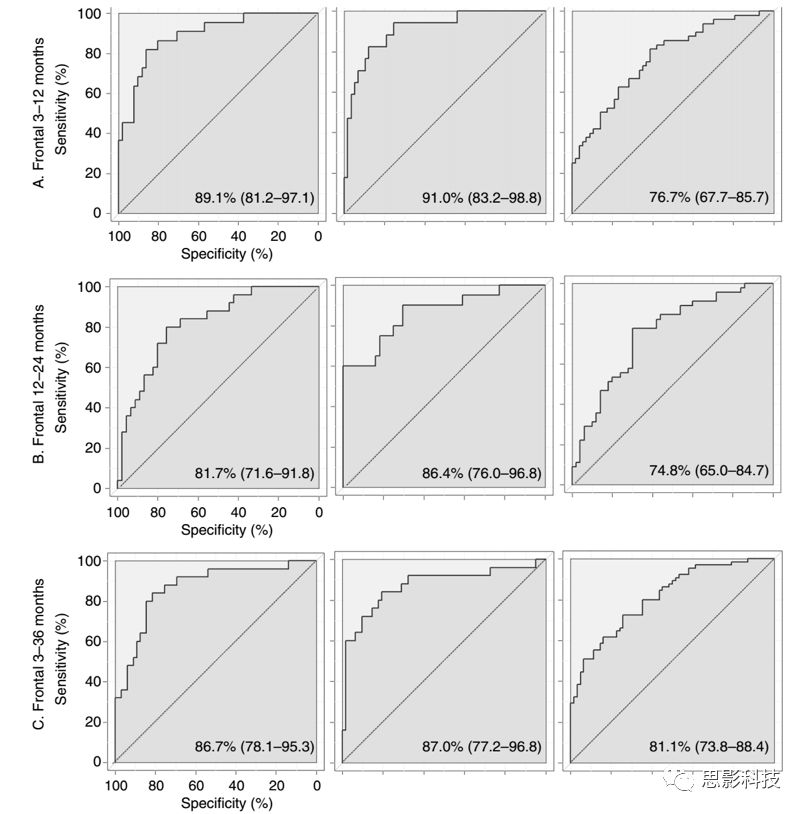
图1 ROC曲线。每个发育阶段每个组别前额EEG模型比较的ROC曲线。每张图的右下角给出了曲线下面积及其95%的置信区间。虚线代表随机分类的准确率,实线代表每个模型的实际准确率
结果
EEG power区分ASD的数据驱动模型
为了确定出生前三年内具体哪个时间段可以最佳区分不同ASD风险的婴儿,研究者构建了一系列线性回归模型。这些模型在3个发育阶段[3–12 months, 12–24 months, and 3–36 months]利用纵向EEG参数区分组别(ASD vs. HRA, ASD vs. LRC, and HRA vs. LRC)。详见图1和表1。对于每个发育阶段每个组别的比较,采用每个感兴趣频段的EEG功率的截距和随年龄变化的发育斜率作为模型参数,选择这两个参数是为了在不同发育阶段能最佳区分不同组别。父母受教育程度和被试性别作为协变量纳入模型。首先,比较不同发育阶段的模型表现以得到最佳区分ASD风险的时间段。同时也利用额叶电极去构建模型并进行比较。最终,比较每个模型选定EEG功率参数和它们的显著性来区分跨时间窗口的纵向功率生物标志。
研究者根据不同发育阶段的ROC曲线来比较数据驱动模型区分组别的性能。值得注意的是,所有模型在95%的置信区间内显著地区分了每组的ROC曲线下面积(AUC)。模型性能的比较基于以下两种情况:在不同发育阶段有效区分ASD组婴儿和其他婴儿、有效区分ASD患病的高低风险。
不同发育阶段ASD诊断结果的鉴别
不同发育阶段和地形分布情况下,3-12个月龄的前额EEG模型能最准确地区分ASD组和其他组(表1)。同时,这些模型特异性更高,预测ASD的真阳性结果(阳性预测值PPV)概率更高,预测ASD的真阴性结果概率更高。初步分析并未发现额叶模型的敏感性高于其他模型,所有模型的敏感性均很高。由于3-36月分组的婴儿数据量更大,因此第二步分析对3-12月龄组所有被试3年内的数据进行了建模,但模型精确度并不如3-12月龄的前额EEG模型。
不同发育阶段ASD患病风险的鉴别
不同发育阶段和地形分布情况下,3-36个月龄的前额EEG模型能最精确的区分婴儿患ASD的高低风险(表1)。同时,这些模型特异性更高,预测真正高风险的比率更高,不过敏感性较低,预测真正低风险的比率更低。在所有发育阶段,风险区分模型的鉴别准确性、敏感性、特异性和真阴性预测率要低于ASD诊断结果。
EEG power生物标志的识别
为了寻找生物标志,比较不同发育阶段不同组别的数据驱动模型的EEG功率参数。模型中特定频段的EEG功率为该频段内所有功率的总和。每个模型中的参数也进行了评估,以确定显著的判别器。模型构建过程中会将被试性别和父母教育程度作为协变量,不过所有模型中,性别差异均不显著,父母受教育程度只有4年大学学历的父母与受过高等教育的父母之间的差异是显著的。大多数情况,是否纳入协变量对结果没什么影响。每个发育阶段内部的比较包含两部分:一是鉴别婴儿是否患有ASD(ASD vs. HRA, ASD vs. LRC),二是鉴别婴儿患ASD的高低风险(i.e., high-risk markers, ASD vs. LRC HRA vs. LRC)。所有个体模型参数的显著性检验都是在多元逻辑回归模型中进行Student’s t tests。
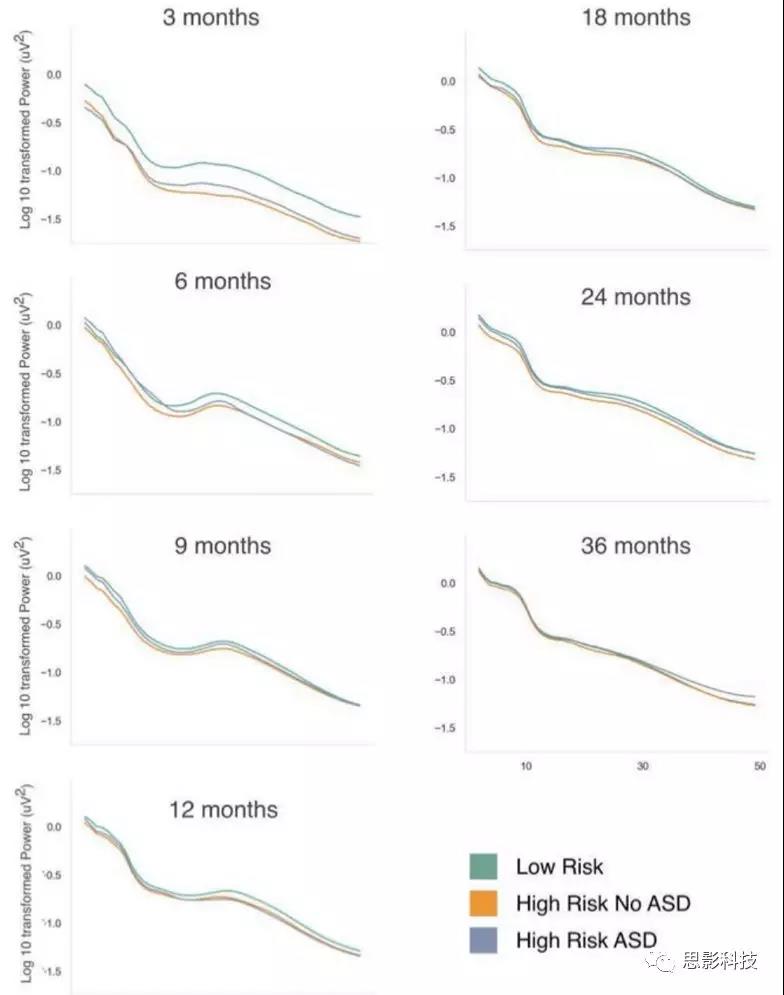
补充图2 前额EEG power随年龄变化的分布
表1 区分病人组的模型表现
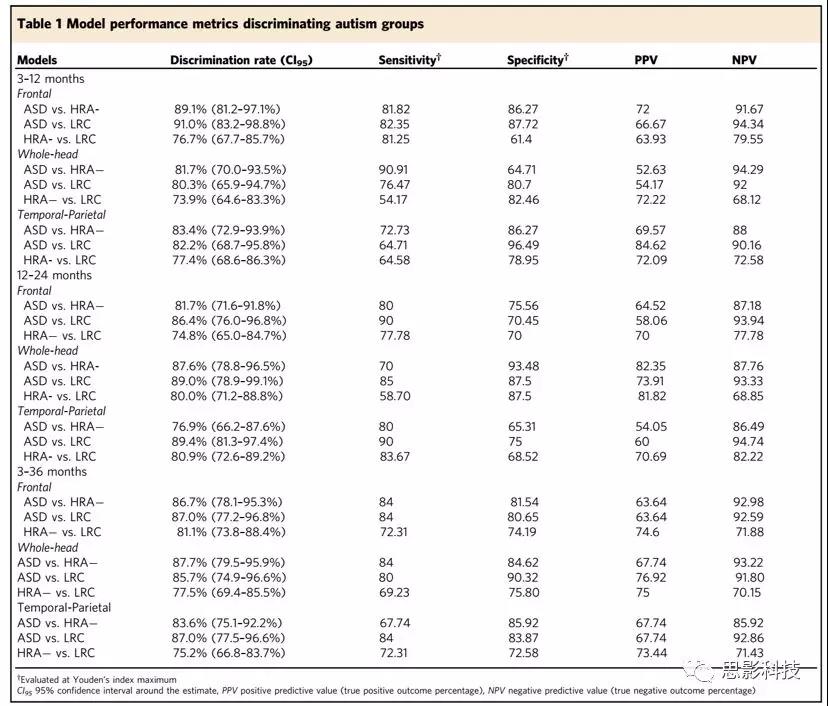
3-12月龄的额叶EEG生物标志
低频段的模型参数能显著区分不同组别(详见表2)。高频段如beta、gamma的模型参数仅能鉴别婴儿是否患有ASD。delta和theta的斜率在识别ASD高风险婴儿时表现优异,表现为较陡峭的delta斜率和较平缓的theta斜率。
表2 3-12个月额叶的power模型
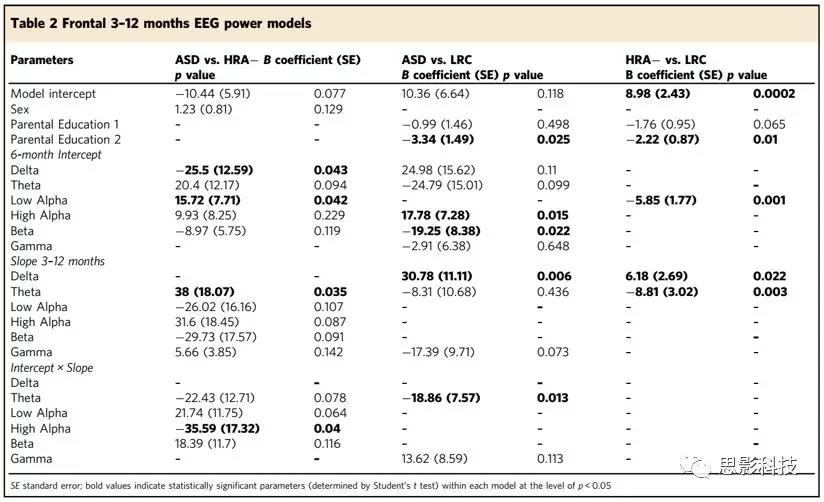
12-24个月龄的额叶EEG生物标志
在第二年的模型中,相比于3-12月龄的模型,显著的EEG参数出现在较高的频率段(详见表3)。仅在区分ASD婴儿和其他婴儿时beta频率是唯一预测变量。12月大时,婴儿较低的beta总功率增加了患ASD的概率。在出生后的第二年,在12个月时,更陡的斜率增加了患高危组ASD的概率,高于其他选定的频带参数。表3 12-24个月额叶的power模型
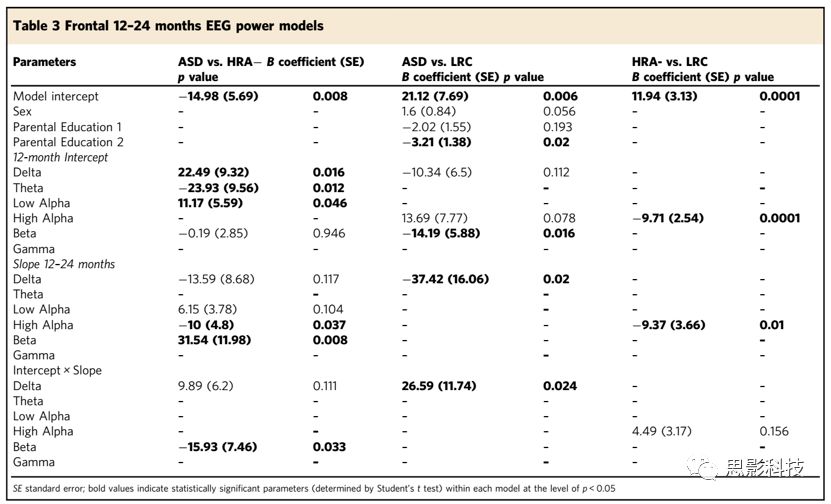
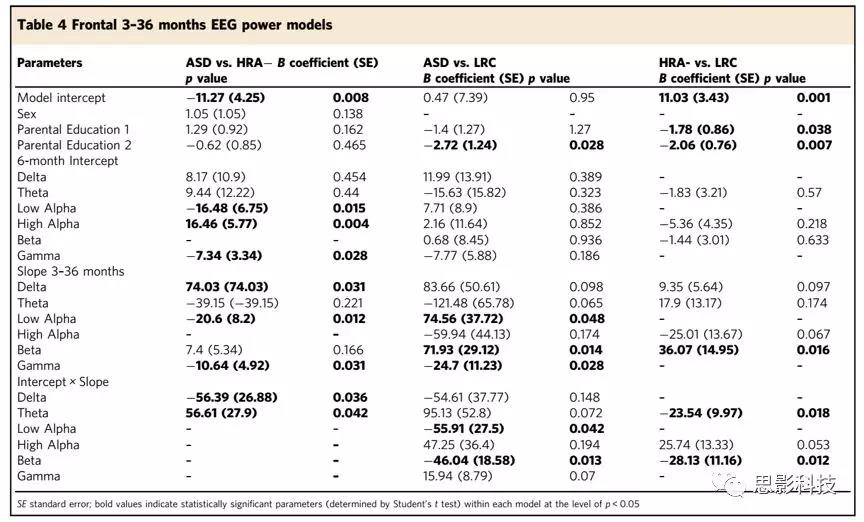
3-36个月龄的额叶EEG生物标志
此时段内,高频段功率更有助于区分不同组别的婴儿。低alpha和gamma的发育斜率是3年内ASD诊断的重要指标。低alpha总功率较小幅度的增加增加了婴儿患ASD的几率(vs. HRA−: slope Student’s t test p = 0.012, vs.LRC: interaction Student’s t test p = 0.042)。gamma总功率较小幅度的改变增加了婴儿患ASD的几率(vs. HRA−, Student’s t test p = 0.031; vs. LRC:Student’s t test p = 0.028)。6个月时的beta总功率和3年内beta总功率的发育斜率存在显著的交互作用,这是将HRA组和LRC组进行区分的重要指标(ASD: Student’s t test p = 0.013, HRA−: Student’s ttest p = 0.012)。
表4 3-36个月婴儿额叶EEG power
讨论
ASD研究尚未解决的一个问题是识别早期的生物标志,以揭示潜在的病理生理学,区分随后的诊断结果。为了应对这一挑战,研究者利用出生后头3年的纵向静息态EEG,这是迄今为止针对ASD低风险家族和高风险家族的婴儿进行的最大的电生理研究。研究者通过关键的发育时间窗口识别EEG Power参数,以识别ASD的患病风险和随后的诊断结果。
研究比较了多个发育窗口和多个地形分布下的EEG power,结果表明出生后第一年的额叶EEG能最好地鉴别ASD的结果。更接近诊断年龄的EEG(行为症状明显时的年龄)并没有为区分结果提供额外的效用,反而在鉴别真正ASD婴儿时出现特异性和能力降低。也就是说,利用EEG power测量对ASD进行归类的能力似乎随着行为测量能力的增长而减弱,EEG测量和行为测量的结果成反比。值得注意的是,尽管男性和女性的ASD患病率不同,但在任何额叶EEG模型中,性别并不是一个显著的预测因素,这表明EEG参数所提供的信息在区分ASD结果方面更为有用。此外,在无ASD婴儿的情况下区分ASD高低风险组的结果表明,早期额叶EEG power足够敏感,足以区分脑功能的亚临床变化。这种区分高低风险组的能力在3年的脑电图测量中确实有所提高,这表明亚临床差异会逐渐出现,或者在更晚的时间出现。
研究结果还表明,EEG power测量的空间定位在区分结果方面很重要,更密集的聚类脑电布局可能比全脑布局有更高的信噪比。值得注意的是,在第一年,额叶区域也比颞顶叶区域提供了更好的分化,这表明EEG power测量的空间特异性对于区分群体是很重要的。在后来的发育窗口中,聚类脑电布局和全脑布局提供了不同的优势,例如额叶和颞叶顶叶布局具有更高的敏感度,而全脑布局具有更高的特异性。因此,脑电通道的空间位置和空间密度是脑电生物标记发育过程中鉴别ASD结果的重要因素,尤其是在出生后早期。
重要的是,研究者确定了不同发育阶段,最佳区分ASD患病风险和后续ASD诊断结果的动力病理生理学。几乎所有的模型都包括来自多个频段的功率参数,这表明跨频段的功率模型参数能更好地区分风险组和患病组。
原文:
Gabard-Durnam, L. J., Wilkinson, C., Kapur, K., Tager-Flusberg, H., Levin,A. R., & Nelson, C. A. (2019). Longitudinal EEG power in the firstpostnatal year differentiates autism outcomes. Nature communications, 10(1),1-12.

微信扫码或者长按选择识别关注思影
第十四届磁共振脑网络数据处理班(重庆,7.26-31)
第三十届磁共振脑影像基础班(南京,7.31-8.5)
第十届脑影像机器学习班(南京,6.30-7.5)
第十二届磁共振弥散张量成像数据处理班(南京,6.18-23)
第二十届脑电数据处理中级班(重庆,8.9-14)
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
小动物磁共振脑影像数据处理班(预报名,南京)
第二十一届脑电数据处理中级班(南京,9.7-12)
第七届眼动数据处理班(南京,7.26-30)
脑电信号数据处理提高班(预报名)
脑磁图(MEG)数据处理学习班(预报名)
思影科技功能磁共振(fMRI)数据处理业务
思影科技弥散加权成像(DWI/dMRI)数据处理
思影科技脑结构磁共振成像数据处理业务(T1)
思影数据处理业务三:ASL数据处理
思影科技脑电机器学习数据处理业务
思影数据处理服务五:近红外脑功能数据处理
思影数据处理服务六:脑磁图(MEG)数据处理
招聘:脑影像数据处理工程师(重庆&南京)
BIOSEMI脑电系统介绍
目镜式功能磁共振刺激系统介绍