

精神疾病的防治是当今世界各国面临的共同难题。在全球范围内,已经造成了极大的经济负担和较高的致死、致残率。据报道,目前我国的精神疾病负担已经超过心脑血管疾病和恶性肿瘤,成为我国医疗体系的最大负担。但是,与其他许多疑难杂症类似(如AD、PD等),精神分裂症的病理性成因至今难以明确。同时,全球范围内,针对精神疾病康复和发病机制的研究也进展缓慢。脑影像数据和基因数据可能是突破其发病机制的关键证据,以往研究已经针对脑影像数据和2万个多个基因点位进行过研究,但到目前为止仍收效甚微。那么,精神疾病的基因表达和其异常的脑结构形态有无关系呢,针对此问题,相关研究者进行了探索性的研究,发现基因SLC39A8的错义突变与壳核灰质体积增大有关。并且这种关系在精神分裂症患者身上明显减弱。该研究发表在著名期刊《JAMA Psychiatry》。
研究意义:
青少年脑发育异常先于许多主要精神症状之前而发生。这种发育轨迹的异常可能与青少年的精神病理的遗传风险有关联。
目标:
本研究的目的是确定与青少年脑结构相关的基因变异,并探索它们二者关联和精神病理的关系。
研究设计和样本:
作者对一组14岁青少年进行了体素水平的全基因组关联研究(GWAS),并利用等位基因特异性表达分析在另外4个独立样本库中的验证作者的结果。将基因与脑的关系在以下三组间进行对比:精神分裂症患者组,未受影响的同胞组和健康对照组。这个基于人口的、结合临床样本的多中心研究纳入了5个中心的样本,分别是:IMAGEN队列,Saguenay Youth Study,Three-City Study, Lieber Institute for Brain Development sample cohorts 以及UK biobank。这些中心的样本同时拥有脑影像和基因测序资料。临床样本包括来自LieberInstitute for Brain Development研究中心的精神分裂症患者和其未患病的同胞。数据分析再2015年10月至2018年4月间进行。
方法:
作者计算了样本的脑灰质容量(脑影像)和基因变异(Illumina BeadChip的基因分型)。
结果:
探索样本包括1721个青少年(873个女孩,50.7%),平均(标准差)年龄未14.44(0.41)岁。重复验证样本包括8690个健康成年人(4497个女人,51.8%),他们来自4个独立的研究。非同义基因变异(SLC39A8中的rs13107325的次要T等位基因,该基因与精神分裂症有关)与壳核灰质体积增大有关(左侧的方差解释为4.21%,t=8.66;95%CI=6.59-10.81,p值为5.35E-18;右侧的方差解释为4.44%,t=8.90,95%,CI=6.75-11.19,p值为6.80E-19)。同时其与壳核的SLC39A8的低表达有关(t=-3.87,p=1.70E-4)。在验证组中这种脑与基因的关系得到了验证,但是精分组(z = −3.05; P = .002; n = 157)和同胞组(z = −2.08; P= .04; n = 149)的脑与基因的关系减弱了。
结论:
作者的结果表明基因SLC39A8错义突变与壳核灰质体积增大有关。这种关系在精神分裂症患者身上明显减弱。这些结果提示异常的离子转运在精神病病因中的作用,并为恢复该突变功能为目的先导干预治疗提供了靶点。
背景介绍
青少年的脑发育经历了结构的改变,如果这个正常的发育轨迹出现了异常可能是许多精神症状的基础。青少年脑发育的生长模式的研究是通过纵向的神经影像研究来实现的,比如尾状核、壳核随成长的减少,或者海马的增加,以及倒U型的杏仁核和丘脑发育轨迹等。双胞胎研究表明,在大脑发育的不同阶段,遗传力(或遗传率)会发生区域特异性的变化,年龄和遗传力的交互作用也会影像灰质体积。青少年大脑结构和患精神疾病风险的共同遗传联系有待去揭示。
GWAS(全基因组关联研究)的meta分析(大规模荟萃分析)是探索脑结构新的基因变异的先进技术。然而,这些研究常常研究的是年龄不一致的人群,从而最大化他们的整体样本量,对青少年脑研究不适用GWAS。因此,人们对遗传因素的了解还很少,无法为我们提供有关大脑发育的正常轨迹的信息,而且与正常轨迹的偏离也被认为与精神疾病的病理生理学有关。为了提高探测青春期大脑中遗传关联的统计效能,很重要就是调查年龄范围较小的样本。这已在2014年的双胞胎研究中得到证实,这项研究纳入了89对年龄相同的双胞胎,并评估了他们的遗传力。他们的统计效能可以与一项大型meta分析相匹敌,这个meta分析的样本是来自不同年龄组1250对双胞胎。使用基于图谱的大脑分割可能会导致检测遗传关联的一些其他缺陷,因为这样定义的大脑区域可能在遗传上是异质性的,因此可能导致假阴性。
为了解决这些局限性,作者研究了2000多名都是14岁的健康青少年(IMAGEN),并将体素级别的脑成像与全基因组关联研究相结合。大脑结构的遗传关联可以出现在特定的发育时期或存在于整个生命周期中。因此,遗传因素可能会导致广泛的神经解剖畸变,这些畸变在特定的发育时期或整个生命周期中都与心理病理学有关。为了验证作者的发现并将作者发现拓展到更广泛的年龄段,作者使用了另外四组健康样本来刻画整个生命周期中的模式。这四个数据库包括Saguenay青年研究(SYS),利伯大脑发展研究所样本(LIBD),英国生物库(UKB)和三城市研究(3C)。对于鉴定出的遗传变异,作者测试了它们对脑组织中邻近基因表达的调控。为了测试青少年大脑的遗传关联是否受到精神病理学的影响,作者比较了患有精神疾病的患者,未受影响的同胞以及健康对照组之间的脑基因关联。
研究方法
研究对象
探索样本以及跨生命周期样本
IMAGEN研究是一项基于人群的纵向影像遗传学队列,招募了2087名14岁的健康青少年,其中1721名参加了vGWAS。作者同时还研究了来自青春期SYS样本的971位健康参与者,来自临床LIBD样本的272位健康参与者,来自基于人群的UKB队列的6932位参与者以及来自基于人群的队列研究3C样本的515位健康的老年参与者。
临床样本
在LIBD的精神分裂症的研究中,作者调查了157例经过治疗的慢性精神分裂症的患者以及149位未受影响的同胞(即他们的兄妹)。IMAGEN项目开展自以下机构:伦敦国王学院,诺丁汉大学,都柏林三一学院,海德堡大学,德累斯顿工业大学,原子能与辅助能源委员会,德国汉堡大学医学中心。对于SYS,所有参与机构的机构审查委员会都批准了本研究报告的所有研究。 LIBD研究作为美国国家心理健康研究所的精神分裂症临床脑疾病分支研究的一部分。该研究得到美国国家心理健康研究所校内计划的机构审查委员会的批准。3C研究得到了比克特医院伦理委员会的批准。每个队列研究发布后,所有成年参与者均提供了书面知情同意书。对于IMAGEN和SYS中的青少年参与者,所有参与者的父母在获得有关研究程序的信息后均提供了书面知情同意,而青少年在获得书面信息后也表示了同意。
方法
全基因组基因型数据
使用Illumina Human610-Quad Beadchip或者Illumina Human660-Quad Beadchip对IMAGEN样本的血样进行基因分型。经过质量控制后,有466114个单核苷酸多态性(SNP)被确定进入以下的分析。有关基因分型和质量控制的详细信息,可在以往发表的文献中,以及在补充方法中的eMethods 1中找到。
结构图像数据
根据针对IMAGEN研究修改的Alzheimer’s Disease Neuroimaging Initiative(ADNI)规程,利用来自3个制造商的3T磁共振扫描仪对研究对象进行了结构磁共振成像。使用SPM8对T1结构像进行了基于体素的形态计量分析(VBM)。预处理步骤包括,分割,空间标准化(MNI空间的Dartel template),调制和平滑化(8 mm FWHM)。
脑表达定量性状位点数据库
在UK Brain Expression Consortium(UKBEC)数据库中,有134个无神经病理学病变的参与者的10个脑区的基因表达数据是可用的。对于基因上任何突变(由vGWAS鉴定),作者首先测试了该SNP是否与该基因的表达相关。其次,作者继续测试这种关联是否是组织特异性的,以及该SNP是否也对邻近(±1 Mb)基因的表达有调控作用。在这样的扩展的搜索中,作者对附近多个基因与大脑多个区域之间的分析进行了多重比较校正(补编中的e方法7)。
统计分析
Voxelwise和全基因组关联研究
在探索样本中,作者对大脑中每个体素的灰质体积进行了GWAS(按照AAL模板标记的438145个体素)。如果一个团块拥有超过217个相连的体素(约4/3×π×[3.3970×1.645] 3 / 1.53像素落入平滑核的90%置信区间内),作者认为该团块具有统计学意义(Bonferroni校正,P <2.4483×10-13=0.05 / 438 145/466114)。然后将已识别的团块视为感兴趣区域,并将每个感兴趣区域的灰质体积由该团块内所有体素的灰质体积的和来计算得到。重复测试也主要是围绕这几个团块展开的。作者通过3000次bootstraps来建立了统计数据的95%置信区间。
基于GWAS汇总数据的孟德尔随机化
对于已经确定的大脑结构,作者通过基于Web的应用程序(MR-Base)进行了基于汇总数据的孟德尔随机化(SMR,MR是基于最基本的孟德尔遗传规律,即亲代的等位基因会随机分配给子代,而基因型决定表型(疾病当然也是一种表型))分析。使用Psychiatric Genomics Consortium 2014 GWAS精神分裂症的结果作为输出,作者测试了确定的大脑结构与精神分裂症之间的关联是否显著,并且排除了非遗传混杂因素。 SMR的显著结果提示暴露(脑容量)和结果(精神分裂症)之间存在关联。这是通过暴露相关遗传变异作为一种手段而达到的,因为遗传变异的随机性质与随机临床试验的设计相仿。尽管SMR的阳性显著结果需要进一步的生物学验证,但阴性的结果至少可以表明缺乏关联。
病人组,同胞组以及健康组的比较
作者首先进行了效应分析,来测试作者是否有足够的样本量来检测作者临床样本中先前确定的遗传关联。为了将精神分裂症患者或同胞与健康对照者之间的关联进行比较,作者使用相关系数估算了其效应量。感兴趣区的灰质体积与SNPs之间的片相关性被确定(控制了年龄,年龄×年龄,性别,智商,总颅内体积以及灰白质体积与总颅内体积之比)。在独立样本之间,作者比较了效应量(即偏相关系数,转换为z统计量后)。95%单侧CI的上限由3000个放回的bootstraps确定,分别用于患者及其匹配的未受影响同胞的2个偏相关分析中中。
结果
人口统计学
在1721名健康青少年的探索样本中(873名女孩[50.7%]),其平均(SD)年龄为14.44(0.41)岁,而8690名健康参与者的重侧样本(其中4497名是女孩[51.8]%])的年龄范围在12至92岁之间。临床样本包括157例精神分裂症患者(其中35例为女性[22.2%],平均[SD]年龄为34.82 [9.91]岁)和149例未受影响的同胞(其中85例为女性[57.1%],平均[SD]年龄为36.60[9.44]岁
精神分裂症风险SNP rs13107325与壳核体积的关系
将基于体素的分析和GWAS(vGWAS)应用于探索样本,作者发现SNP rs13107325的次要T等位基因(基因SLC39A8中的错义突变)与双侧壳核的体积增大相关(左半球:t1705 = 8.66;P = 5.35× 10-18;方差解释度为[VE] = 4.21%;右半球:t1705 = 8.90;P = 6.80×10-19;VE = 4.44%),并且这些团块在左右半球之间是不对称的(图1A-C)。此外,作者发现SNP rs7182018的minor Gallele(lncRNA RP11-624L4.1的内含子变体)与双侧中央沟(左半球:t1705 = 9.86; P = 1.25×10−22; VE = 5.39%; 右半球: t1705 = 9.96; P = 4.54×10−23; VE = 5.50%; 图1D和E;)的2个团块的灰质体积增大相关。
rs13107325与精神分裂症相关(2014年PsychiatricGenomics
Consortium [phase 2] GWAS)。SMR结果确定了壳核的灰质体积与精神分裂症之间的关联(左壳状核:b = 0.9388; SE =
0.1329; P = 1.61×10-12;右壳核簇:b = 3.444; SE =
0.4875; P = 1.607×10-12)。考虑到SMR分析未发现中央沟与精神分裂症之间存在关联,作者推测rs7182018与精神分裂症无关(图3)。
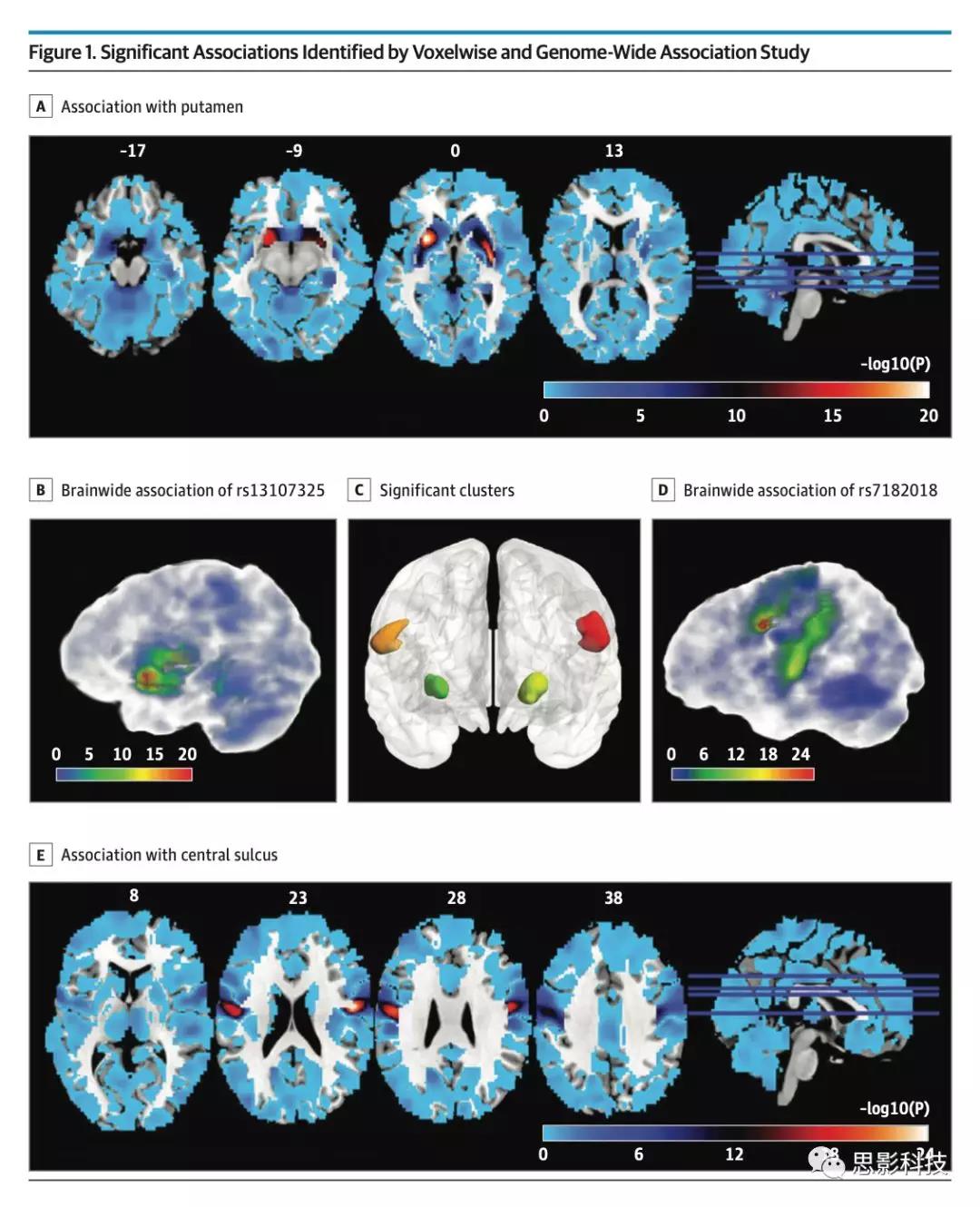
图1. 体素和GWAS关联
注释:大脑每个体素的灰质体积与SNP rs13107325(A和C)或rs7182018(D和E)之间的显著关联(-log10 P)。红色表示较强的关联,而蓝色表示较弱的关联。B,经过Bonferroni校正的四个团块(P <2.45×10-13,由0.05 / 466114 [SNP数] / 438 145 [体素数]计算得到)。左右中央沟周围的两个团块分别用红色和橙色标记。左右壳核分别用黄色和绿色标记。
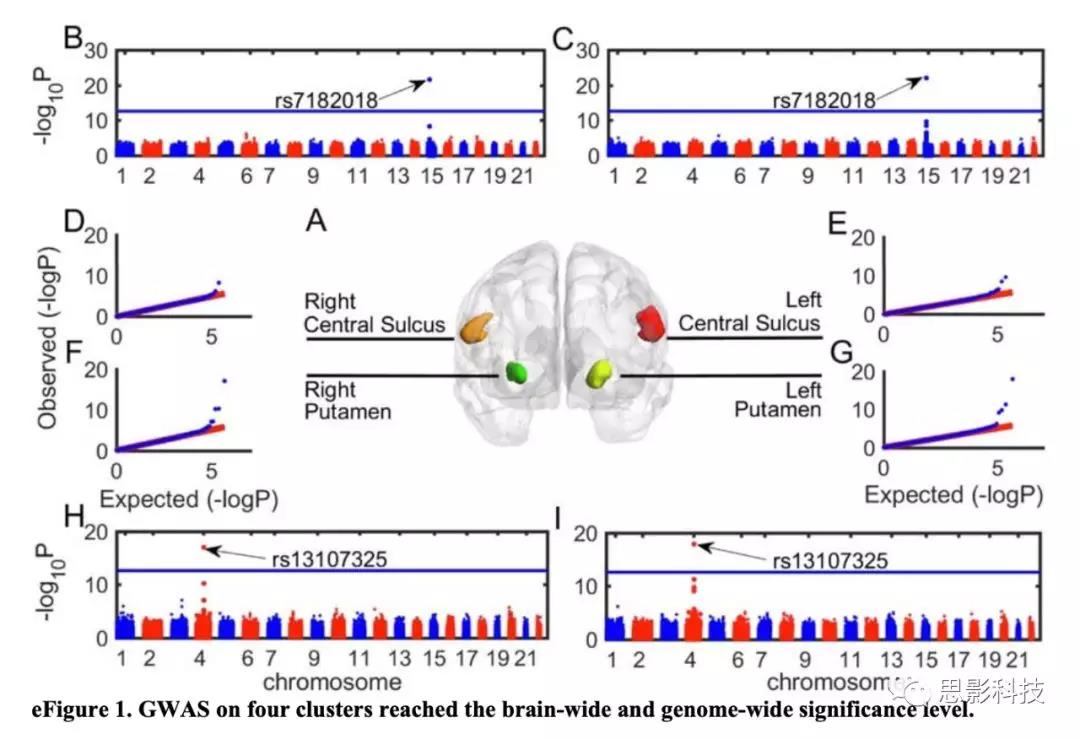
图2. 四个GWAS和关联的体素与基因表达的具体关系
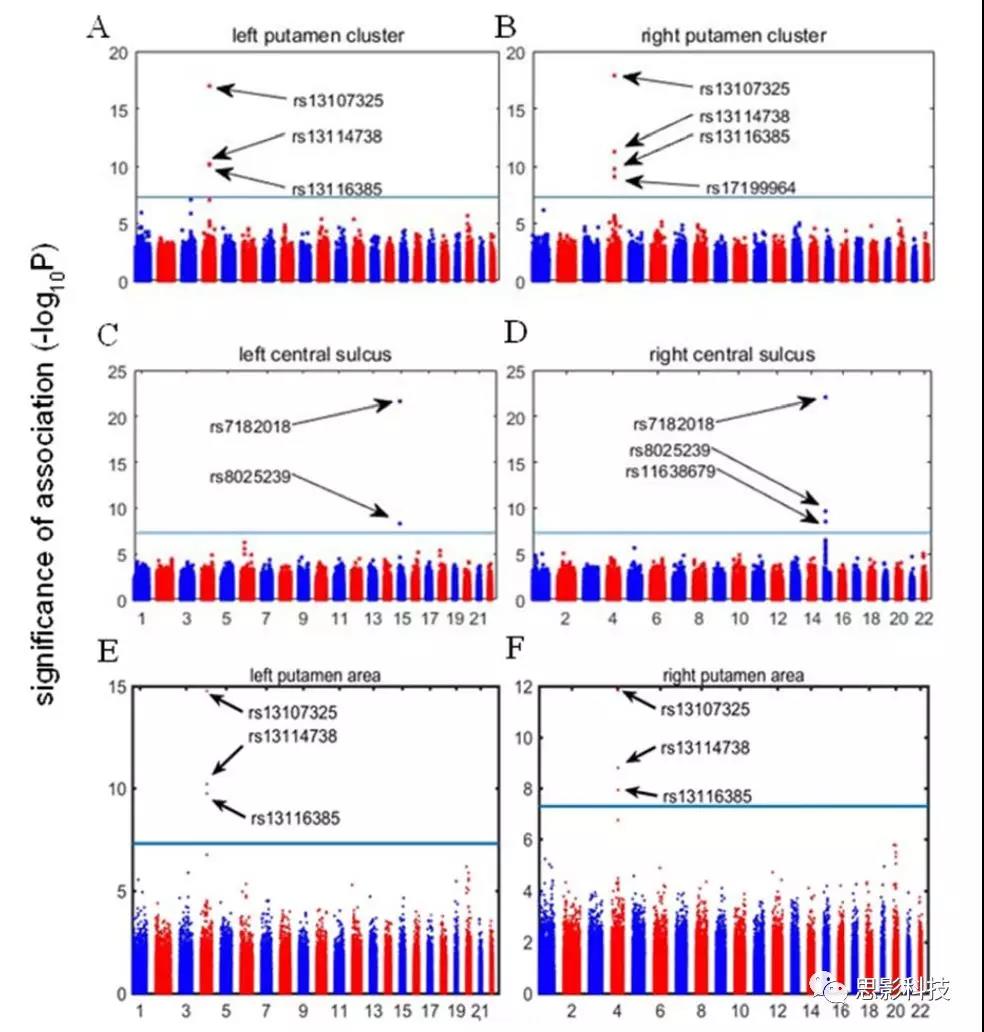
图3. 全脑探索性分析发现的与GWAS相关的中央沟和壳核相关的基因表达
跨生命周期的独立样本重测
在971名平均(SD)年龄为15.03(1.84)岁的健康青少年的SYS样本中,作者在左侧壳核中复制了探索样本中发现的SNP rs13107325正相关关系(t964 = 3.70; P = 1.16×10−4),但没有发现右侧壳核中的这种关联(t964 = -1.73; P = .08)。右侧壳核的团块显著受到SYS样本中岛叶皮质的较大变化的影响:一部分岛叶皮质被映射到该团块中。
使用UKB样本(平均[SD]年龄为62.64 [7.41]年;n = 6932),作者重复了rs13107325与壳核灰质的正相关关系(左半球:t6885 = 4.80; P = 8.16×10-7 ;VE = 0.33%;右半球:t6885= 4.80;P = 8.16×10-7;VE = 0.60%)。由于该队列的样本量很大,作者进一步证实一些有显著意义的团块,(通过在团块水平上,将SNP应用于全脑,并进行10,000次的置换检验)。在另外两个平均年龄(SD)为31.92(9.50)的独立样本中(LIBD样本,n = 272)和77.48(5.12)(3C样本,n = 515),作者再次确认了前面发现的正相关关系。
rs13107325与壳核SLC39A8低表达的关系
使用UKBEC(n = 134、112CC基因型,22 CT CT基因型,SNPrs13107325的0TT)的表达定量性状基因座(eQTL)数据库,作者发现风险等位基因(T)atrs13107325的携带者表现出SLC39A8的较低表达(t127 = -3.87; 95) CI(%6.5--1.73; P = .0002)(图2A和B)。此外,作者发现尽管SLC39A8(图2C)在脑内广泛表达,但QT缔合对壳核是特异性的,在任何情况下均未检测到其他大脑区域(P <.0008,对10种类型的脑组织和6个邻近基因进行Bonferroni校正)(图2D)。除了基因SLC39A8(附录中的表13和表14),作者还发现rs13107325与海马中NF-κB1的较低基因表达有关(t120 = -3.62; 95%CI,−6.31至−1.28; P =)。 0004),额叶皮层中的MANBA(t125 = -3.73; 95%CI,-5.93至-1.84; P = .0003),CENPE在枕叶皮层中的表达较高(t127 = 3.69; 95%CI 1.72至6。 10; P = .0003)。
精神分裂症患者风险基因导致基因-脑关联减弱
尽管精神分裂症中对于壳核的结构神经影像学结果不一致(无差异,减少或扩大),但长期以来,这种结构与精神分裂症中多巴胺合成能力的提高和额纹状体连接紊乱有关,并且是各种抗精神病药物关键靶点。为减少干扰因素,作者使用了未受影响的同胞(其患有精神分裂症的遗传风险较高,但没有临床表现和治疗效应的患者)来进一步验证了精神分裂症中rs13107325与壳核的关联。
作者假设与健康对照组相比,rs13107325与壳核的关联在患者和未患病的同胞中均明显减弱。由于健康对照大的效应量(r = 0.3117; n = 272),效应分析估计95%CI,以及显著性水平时的样本量为102(单尾)。因此,LIBD研究中的患者(n = 157)和未患病的同胞(n = 149)有着足够的样本来检测这种关联。作者发现右半球中的rs13107325-壳核关联在患者和未受影响的同胞中没有统计学意义。这种异常可能是特异的,因为rs7182018-中央执行网络关联在所有3组中均保持显著。与健康对照组相比,患者的rs13107325-壳核关联显著减弱(z = -3.05; P = .002)。个人(z =-2.08; P = .04)。接下来,作者证实同胞中的关联也减弱了。同时,病人组比同胞的这种关联更弱。
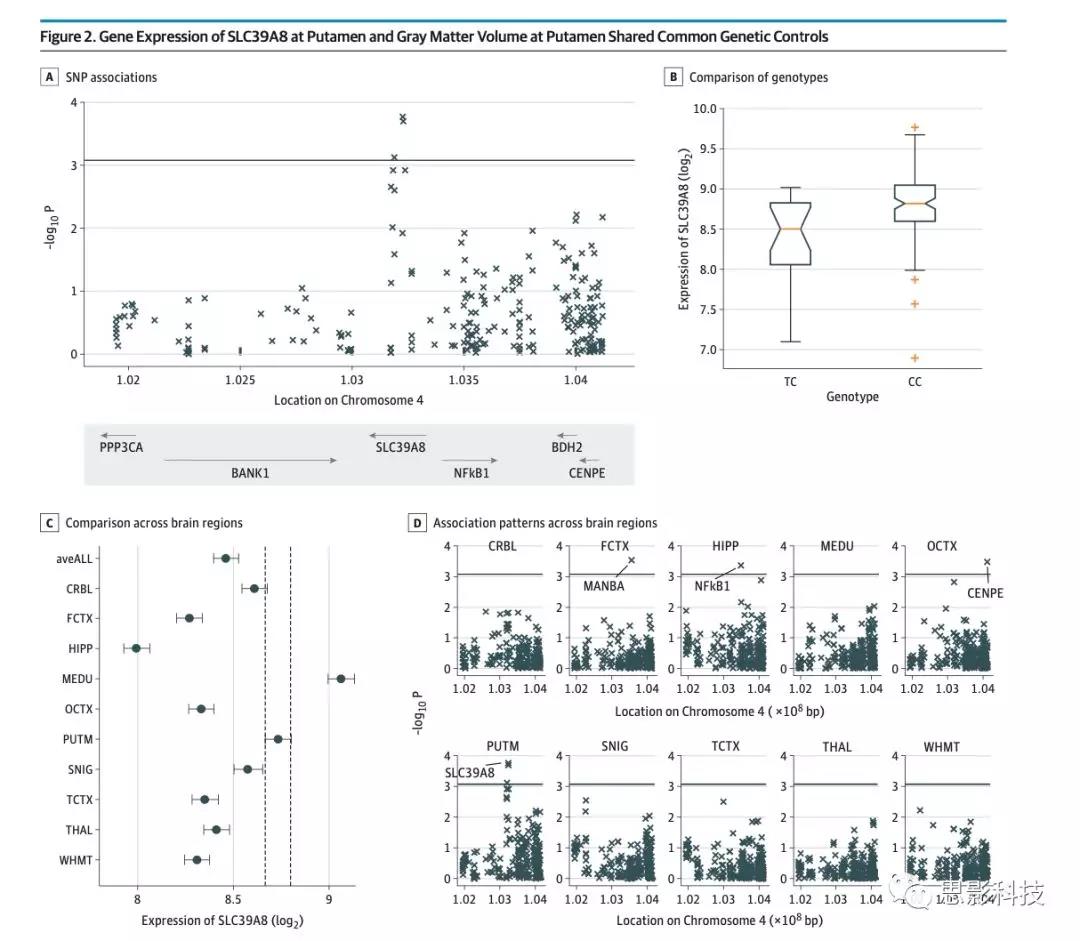
图2.壳核SLC39A8的基因表达和拥有共同基因控制的壳核灰质体积
A, 单核苷酸多态性(SNP)rs13107325与SLC39A8邻近基因的基因表达水平之间的关联的统计学差异意义水平(Affymetrix探针是根据其起始碱基在4号染色体上的位置进行组织的)。
B, SNP rs13107325不同基因型的壳核SLC39A8基因表达水平之间的比较。
C, 10个脑区的SLC39A8的基因表达水平(均值和95%置信区间)的比较。
D,SNP rs13107325与10个脑区的基因表达之间的关联模式。具有显著关联的基因(P <.0008,通过Bonferroni校正:0.05 / 10/6)用基因名称标记。 bp表示碱基对。
讨论
在此vGWAS中,作者在青少年脑组织中发现了rs13107325和壳核的关联,并在整个生命周期中证实了这种关联。孟德尔随机分析显示,核壳灰质体积与无非遗传混杂因素的精神分裂症之间存在显著相关性。在患者的未受影响同胞中,发现rs13107325-壳核关联明显减弱,这可能是由于精神分裂症的遗传风险所致。总之,这些发现为精神分裂症的病理学以及决定壳核灰质体积的机制之间存在相互作用提供了新的且可以验证的假设。
单核苷酸多态性rs13107325(位于SLC39A8的外显子,染色体4)编码在质膜和线粒体中表达的溶质载体转运蛋白ZIP8。 SLC39A8跟精神分裂症有关联,这已通过大规模GWAS和全基因组DNA甲基化分析(从24名患者和24名正常被试脑中收集)得到证实。自2012年以来,学术界认为该基因可能参与了精神分裂症的精神病理,并已证明其涉及免疫过程,谷氨酸能神经传递和大脑中必需金属的平衡调节。
有人推测SLC39A8与精神分裂症之间的关联可能与其在大脑发育过程中参与促炎性免疫反应有关。作者的发现强调了SLC39A8对核因子-κB(NFκB)通路的负调节是一种可能的因果机制。NFκB通路诱导了与精神分裂症症状相关的促炎基因(例如细胞因子)的表达。在健康人群中,SLC39A8与壳核灰质体积之间的强关联可能与NFκB在大脑发育过程中对神经元生长和神经元形态的调节作用有关。在精神分裂症患者中,这种减弱的关联可能是由于基因和蛋白质水平上的NFκB失调以及患者脑组织中的细胞核激活所致。rs13107325是一种错义突变,将丙氨酸(非极性)替换为甲腺原氨酸(极性)(Ala391Thy),导致ZIP8-Thy391将更少的金属离子转运到细胞中。因此,在由大样本GWAS发现与精神分裂症风险有关的SNP rs13107325之后,作者的结果表明精神分裂症的分子病理学可能会破坏壳核发育过程中的神经元离子介导的调控。
IMAGEN样本的1721个14岁健康青少年提供的效应大小比UKB样本(6932)大3倍。遗传因素可以解释多达80%的脑部解剖学的遗传,其中多达54%可以被大量的SNP解释。然而,根据以往文献,由单个遗传变异解释的方差百分比仅为0.52%。在这项研究中,鉴定出的遗传变异解释了所观察到的灰质体积变化的4%以上。对青少年大脑的如此大的单变量遗传影响可能是由于在以往环境对其的累积影响较小。对同龄青少年的分析也许可以帮助作者解释为什么这种新颖的关联未能被以前纳入不同年龄组的大规模meta分析所识别。
结论
作者使用新颖的方法鉴定了一个新基因,该基因同时与离子转运蛋白和免疫反应相关,他们都与精神分裂症发展有关的。鉴于SLC39A8基因的主要功能可通过药物操作而获得,作者认为这些结果对于发现精神分裂症的新疗法至关重要。
原文:
Association of a Schizophrenia-Risk Nonsynonymous Variant With Putamen Volume in Adolescents: A Voxelwise and Genome-Wide Association Study
Q Luo, Q Chen, W Wang, S Desrivières… - JAMA …, 2019 - jamanetwork.com

微信扫码或者长按选择识别关注思影
第十四届磁共振脑网络数据处理班(重庆,7.26-31)
第三十届磁共振脑影像基础班(南京,7.31-8.5)
第十届脑影像机器学习班(南京,6.30-7.5)
第十二届磁共振弥散张量成像数据处理班(南京,6.18-23)
第二十届脑电数据处理中级班(重庆,8.9-14)
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
小动物磁共振脑影像数据处理班(预报名,南京)
第二十一届脑电数据处理中级班(南京,9.7-12)
第七届眼动数据处理班(南京,7.26-30)
脑电信号数据处理提高班(预报名)
脑磁图(MEG)数据处理学习班(预报名)
思影科技功能磁共振(fMRI)数据处理业务
思影科技弥散加权成像(DWI/dMRI)数据处理
思影科技脑结构磁共振成像数据处理业务(T1)
思影数据处理业务三:ASL数据处理
思影科技脑电机器学习数据处理业务
思影数据处理服务五:近红外脑功能数据处理
思影数据处理服务六:脑磁图(MEG)数据处理
招聘:脑影像数据处理工程师(重庆&南京)
BIOSEMI脑电系统介绍
目镜式功能磁共振刺激系统介绍