

本项研究通过记录左半球视觉区域内2分钟电生理反应的方法,在5岁学龄前儿童群体中发现了稳定且独特的对文本信息敏感的脑电信号。这一脑电信号与学龄前儿童的基本字符认知能力(一项独立采取的行为测量)有显著相关性,说明存在除了视觉熟悉之外的特异化神经回路,这些发现还强调了高度灵敏客观的非行为测量方法对发展中个体字符认知能力(阅读能力的前身)评估的潜力。本研究由比利时鲁汶大学心理科学研究所和神经科学研究所的Aliette Lochya, Marie VanReybroecka, 和Bruno Rossion发表在PNAS上。
摘要
阅读是人类社会最重要的文化发明之一,熟练的成人读者很大程度上依赖于左半球后脑区进行阅读加工。对儿童群体研究通常发现在接受了约1 年的正式教育和阅读习得之后,字符串左半球皮层特异化。
本研究中,我们用每5个快速呈现的伪字体中插入一个字符串(每秒6个项目)的范式向5岁(n=40)前阅读者呈现刺激,并记录儿童的头皮电生理反应(EEG)。2分钟的记录发现,字符串在左侧枕颞皮层上诱发一种强烈的与1.2hz(即6hz/5)的预定频率有关的特异性反应。这种电生理反应幅度的个体间差异与被试字符认知显著相关,字符认知是儿童日后阅读能力的一个预测因素。这些结果表明,这种快速记录的非行为测量方法在评估发展中人群的阅读能力方面具有很高的潜力。
第二个实验(26名学龄前儿童)中验证这些发现,在第二个实验中,熟悉的符号和物体的线条分别诱发了大脑右侧和双侧特异性反应,显示了早期字符串左半球优势加工的特异性。总之,这些发现表明,在接受正规教育之前,儿童有限的文本知识足以发展出专门的左半球偏侧化神经回路,进而表明重复性声音在左半球的加工对左半球后脑区发展的影响出现在早期并发展迅速。
关键词: EEG 字符串 儿童皮层特异化 左半球
前言
精通阅读的成年人主要依赖左侧颞枕腹侧通路加工阅读这项复杂却快速有效的认知工作。在儿童阅读习得时期,通过声音重复映射到同侧大脑后部字符串表征区逐渐形成左半球特异性(左侧颞叶和额叶区域表征)。尽管这种左侧化的确切开始时间并不为人所知,但在正式的阅读习得过程中,这一过程似乎要花费相当多的时间。
功能磁共振成像(fMRI)研究发现典型字符串的偏左侧化的大脑模式仅出现在阅读精通者,而自然环境中,显著的左半球偏侧化只出现在约7岁已经具备正式阅读能力的儿童上。在直接测量大脑活动(EEG/MEG)的发展研究中,字符串出现后大约200ms诱发正常阅读成人颞枕区域激活(比如,N1,N170), 而该成分未出现在学龄前儿童组,即使这些儿童已经可以成功分类或命名字符串。一般情况下,在1年或1.5年学龄后,儿童逐渐对文本敏感。而且,经过短音素训练的学前儿童对字符串可以产生N1效应,但是这种反应既不是左半球偏侧特异性也不持久,训练中断几天后就会消失。总的来说,这些发现和少量学龄前儿童数据共同说明:字符串左半球偏侧特异性仅在正式学习或强化训练后出现。也就是说,对于文本左半球后部偏侧化加工来说,较高程度的字符-语音自动化关联显得很必要。
文本材料在左半球皮层特异化的明显迟发是相当令人惊讶的,因为学龄前儿童通常已经获得一些字符认知,这种认知能力是随后阅读能力的最重要预测因素之一。因此,如果字符认知不仅是感觉相关的,而且还涉及到图形-音位关联,那么文本的神经特异化通路将比通常想象的更早和/或更快开始。不幸的是,功能磁共振成像不太适合幼龄(学龄前)儿童,这使得阐明左半球皮层特异化的开始时间变得困难。至于标准的EEG方法,他们需要许多试次来达到可接受的信噪比水平(SNR),因此,可能无法在短时间的记录过程中检测出早期幼儿左侧大脑皮层的这种特殊性。这种研究方法将产生重要的理论和社会后果,因为它将揭示自然设定条件下基本字符认知足以诱发皮层特异化。此外,它还可以开辟新的视角来评估早期和整个发展阶段的阅读加工。
本研究通过最近发展的oddball范式,探索学龄前儿童早期文本大脑左半球偏侧特异化的假设(实验1:n=40,平均年龄=5.5;实验2:n=26,平均年龄=5.8)。该范式将偏差刺激(“oddball”)周期性地插入到快速周期性呈现的标准刺激(“base”)中(图1和视频S1)。与标准的事件相关电位(ERP)测量方法相比,这种快速周期性视觉刺激(FPVS)方法具有很高的灵敏度(即高信噪比)。最近的一项研究发现每个成人参与者的左半球后侧对单词和对照字符(伪字体,pseudofonts)加工存在显著差异。
本研究在实验1中,我们设置2种对比条件:以每秒6项的速度,在伪字体(pseudofonts)中周期性(1/5项)插入字符串(字或伪字,words or pseudowords),以及在伪单词中插入的单词(图1)。
在实验2中,我们设置2类任务:字符串辨别,高对比度/空间频率但视觉熟悉度不同的刺激辨别:熟悉的键盘符号(familiar keyboard symbols)与伪符号(pseudosymbols),陌生的外来文本(unfamiliar foreign script)与伪文本(pseudoscript),以及物体线条图(line drawings of objects)与伪图形(pseudodrawings)。在所有条件下,如果被试能将新异刺激与标准刺激区分开来,则预期在精确的1.2Hz(即6/5Hz)及其谐波(如2.4Hz)出现反应,从而在脑电频谱中提供视觉辨别的客观(即实验上预先定义的)证据。
用这种方法对在正式阅读习得之前的学龄前儿童进行测试,提供了一个独特的机会来评估字符串的早期左半球皮层特异化(假定)与前阅读能力之间的关系。尽管早期的左半球偏侧化可能是由于低水平的视觉特征敏感性(例如,高空间频率)引发,但它与字符认知的个体差异以及与声音无关的熟悉符号的双侧或右偏侧化(如实验2所测试的)相关,进而支持在整个发展过程中语音重入(reentrant)映射加工出现较早且快速发展的观点,具有重要的理论和社会意义。
实验材料与方法
被试
实验一:43名幼儿园三年级儿童(23名男性,平均年龄=5.48岁;年龄范围=5.01-5.94岁)参与实验,其中3名被试数据排除在外(见下文)。视力正常/矫正正常,并在家长知情同意,鲁汶大学生物医学伦理委员会批准后进行测试。儿童不知道这项研究的目的,在刺激呈现过程中,刺激类型以周期性的速度发生变化。
实验二:26名儿童(14名男性,平均年龄=5.82岁;范围=5.13-6.09)进行了测试。
行为实验
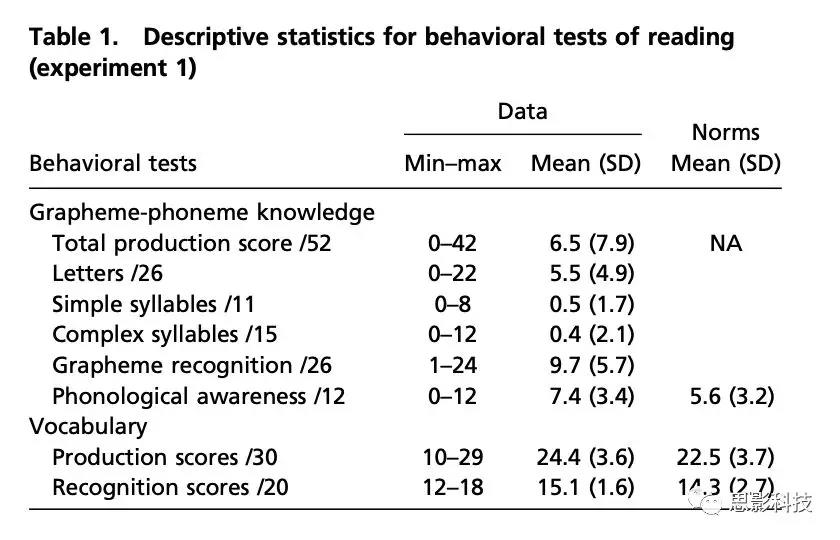
实验1:孩子们接受了WISC(立方体和符号搜索),言语广度和词汇(ELO;27)筛查测试。在此基础上,排除了一名在多项任务(词汇:Z=-3.38,语言广度:Z=-2.57,WISC子测验(标准注释:1和4))中得分异常低的儿童(图S3)。对于与阅读相关的测量,我们测试了音素意识(初始音素识别,N-EEL;26)、字形识别(通过指认)和字形音素产出(即字符阅读),阅读简单和复杂的音节(BELO;46)(表1)。字符用小写字符呈现。字符名称和发音都被认为是正确的。计算Z值以去除当前样本分布中的极端值。一个孩子的数据被删除(字形-因素产出得分:Z=+4.48)。试验及结果详见表1。 表1 实验2 测试儿童的视觉注意能力、词汇、字形音素产生和识别能力
EEG实验刺激
实验1:三类15种刺激(单词、假单词、假字 (words,pseudowords, pseudofonts))由四个符号项(字符或假字(lettersor pseudofont))组成。在第一种情况下,“标准”刺激是伪字体,“新异”刺激是单词(PF-W)(图1)。在第二种情况下,在伪字体中插入新异的伪词(PF-PW)。这个条件被用来排除语义处理对单词和伪字体之间假设的左偏判别反应的任何潜在贡献。在最后的条件下,将新异单词插入到作为基础刺激的伪文本中(PW-W),使得能够评估正字法的词义检索。法语单词[常见名词(估计频率:242.01/million)单数形式]选自Manulex数据库。伪单词是通过重新排列单词的第一个和最后一个字符(相邻字符(adjacent letters))而构建的(例如,“joli”(“可爱”)和“fète”(“一方”)这两个单词可能会产生伪单词jote和fèli)。单词(5.586,SD:927.48)和伪单词(6.278,SD:924)的平均连接单元频率没有差异[F(1,28)<1;P=0.6]。假词在法语中是可发音的,并遵守语音规则。逐项构建伪字,垂直翻转每个单词,并通过Adobe Photoshop分割成简单的特征。然后,根据字符总数(4)和原始单词的总大小(宽度×高度)(24),重新排列这些片段以形成伪字符。因此,伪字符包含连接、上升/下降特征和特写形状(图1为刺激示例)。因此,每个词都有相应的PW和PF,包含数量完全相同的黑白对比,因此所有条件在低级视觉特性方面都是可以对比的。
实验二:为每一类项目建立了15个刺激,以及相应匹配的15个视觉对照刺激。在所有条件下,视觉对照项的构建方式与实验1中描述的完全相同,因此每个新异刺激都有一对由其重新排列的片段组成的视觉对照刺激。在PF-PW条件下,用4个字符的15个假词作为新异刺激。在第二种情况下,键盘符号(&V§!% ? =+<>*)被排列成四个元素序列,其中允许序列中相同符号重复(以模拟单词或伪单词中相同字符的重复)。在第三种情况下,选择Kartika、Cyrillic和希腊字体(文字处理器)的元素,随机混合并排列成四个元素序列。最后,从“Wingdings”字体中选择线条(line drawings)(排除黑色的形状、面和同一形状的变体)。在这两个实验中,刺激物在阴极射线管(CRT)显示器(屏幕分辨率为800×600,刷新率为60hz)上以Verdana字体(script和线条除外)集中呈现。图像的高度在40到78像素之间,宽度在92到184像素之间,这取决于单个字符的形状。在1米的视距下,刺激视角范围为3.1到6.3(宽度)和1.1到2.2(高度)。
实验程序
每个刺激序列十字形注视点呈现2-5秒,2秒后注视点逐渐消失,40秒刺激序列,2秒刺激逐渐消失顺序呈现。在40s内以6hz正弦波规律呈现刺激,83ms(即一个周期持续时间=166ms)后所有刺激完成一个呈现周期(图1)。考虑到刺激可以在低对比度(即20%或更低)下识别,刺激可见性的实际持续时间接近140ms。每五分之一序列(1.2hz新异频率,833ms)项是一个单词(PF-W和PW-W序列)或伪单词(PF-PW序列)。用Matlab7.8(Mathworks)和PsychToolbox(48;见PsychToolbox.org/)呈现刺激。由于该方法信噪比较高并快速呈现刺激,在实验1中,每个条件仅使用两个40s的刺激序列,因为本研究的目标之一是在最短的时间内获得对视觉词汇的选择性反应。实验一总共40s(序列持续时间)×2(重复次数)×3(条件)设计,共4min刺激呈现。在实验2中,每个序列被呈现3次,总共40s(序列持续时间)×3(重复)×4(条件),共8min刺激呈现。在每个刺激序列之间有大约30秒的暂停,这是手动设置以确保低伪影EEG信号。
在视觉刺激呈现过程中,儿童注意十字形注视点,注视点出现任何短暂(200毫秒)颜色变化(蓝色到红色;每个序列随机6次变化)时按空格键。这个独立任务被用来在整个实验过程中保持恒定的注意力水平。儿童几乎以91-95%正确率完成这项任务,表现出对刺激的高度注意,并且在反应时间上没有条件差异(PW-W:650 ms;PF-PW:665ms;PF-W:672 ms)(F(2,62)<1)。
EEG采集和预处理
在学校安静的房间里,儿童舒适地坐在离电脑屏幕1米的地方。使用32通道Biosemi Active II系统采集EEG信号,采样率1024hz,电极位于标准的10-20个系统位置。中线位置电极为参考电极(common mode sense),所有电极的波幅保持在50毫伏以下。所有脑电图分析均在Letswave 5和Matlab2012(Mathworks)进行。0.1-100 Hz快速傅立叶变换(FFT)带通滤波之后,EEG数据以包括每个序列之前和之后的2 s的时间窗分段(-2~42 s)。然后将数据文件重新采样至250Hz,以减少文件大小和数据处理时间。用线性插值法代替伪影或噪声信道。所有通道都以全脑共同平均值进行重参考。然后,脑电信号从刺激开始到39.996s进行再次分割,与刺激中48个完整的1.2hz周期完全对应。该持续时间对应于刺激周期40 s内833ms新异刺激(1.2hz)的最大完整周期。
频域分析
为了提高信噪比,在时域内对每个参与者的每个条件下所有试次进行平均。对平均时间窗进行FFT,提取各电极的标准化波幅,产生了高频分辨率(1/39.996s=0.025hz)的脑电频谱,增加了信噪比,且可以准确识别感兴趣的频率(即,6hz为标准刺激频率,1.2hz及其谐波为新异刺激频率)反应。为了估计整个脑电频谱的信噪比,将每个频率点处的波幅除以周围20个频率点(每侧10个)的平均波幅。为了量化感兴趣的反应,感兴趣的频率点减去20个周边频率点(即噪声)的平均波幅。基于每种情况下的总平均波幅谱,计算每个通道的Z分数,以评估是否存在明显的新异刺激诱发的频率和谐波反应以及标准刺激诱发的频率和谐波反应。Z值大于2.58(P<0.005,单尾,信号>噪声)被认为是显著的。像其他成人的字符串研究或类似的面部研究一样,使用保守阈值是因为所有通道的反应及其在不同谐波上的表现都进行了评估(尽管我们预期反应出现在大脑后部)。基于任何电极上最大数量的连续显著的谐波(#4),在所有条件和电极上选择相同数量的谐波。最后,为了量化多个谐波的周期性反应,对每个参与者减去基线波幅的显著谐波(不包括标准刺激频率)求和。一名儿童的数据被删除,因为他的反应(3.92μV)高于该组平均反应(1.25μV,SD=1.2μV)的2个标准差。
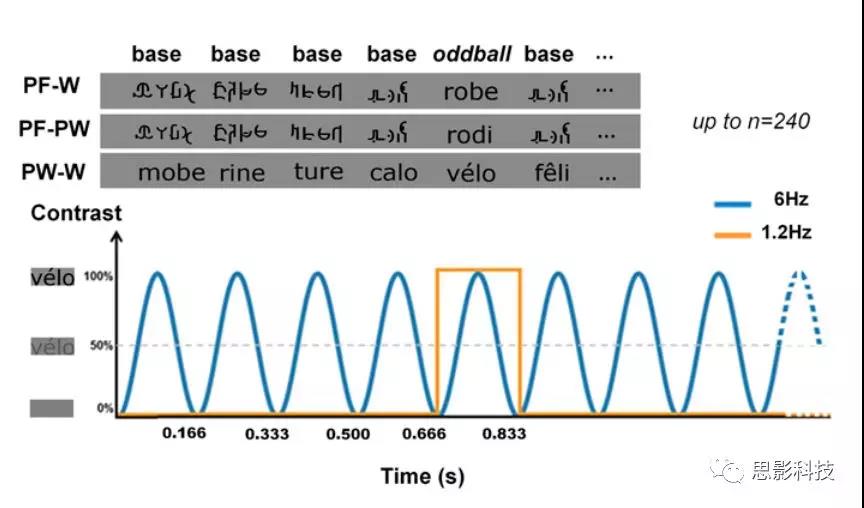
图1。实验范式(实验1)。每种条件下,两个序列共有96个新异刺激。在40s内以6hz正弦波规律呈现刺激,83ms(即一个周期持续时间=166ms)后所有刺激完成一个呈现周期。刺激在标准刺激(b)和新异刺激(o)之间交替,例如:bbbbobbbbobb。标准刺激要么是伪字(顶部和中部),要么是伪单词(底部),新异刺激要么是单词(顶部和底部),要么是伪单词(中部),每五项出现一次(6/5hz=1.2hz)。刺激非立即重复随机呈现。
结果
oddball 反应
对于插入伪字体的字和伪字,地形图和脑电频谱结果发现左侧枕叶O1在1.2hz(一次谐波),2.4Hz(二次谐波)和3.6Hz(三次谐波)出现明显的(即信噪比>1.6,信号增幅>60%,基线校正幅度之和>1.4μV)辨别反应(图2A)。在这两种条件下,在大脑后部七个电极(O1、O2、Oz、P7、P8、PO3、PO4)上,从1.2Hz到4.8Hz(F/5–4F/5)一个或多个谐波均表现出显著反应。然而,与成人加工模式不同,在伪词序列中插入的词没有呈现辨别反应(图2)。所有电极均根据其最大波幅值进行排序,该值为基线校正后四次谐波(1.2 Hz、2.4 Hz、3.6 Hz、4.8 Hz,所有P值<0.005)的总平均(F/5–4F/5;见材料和方法)。伪字(PF)和伪词(PW)条件下的最大反应出现在O1。
基于这一结果,在实验1中,用3(实验条件:PF-W,PF-PW,PW-W)×2(半球:O1,O2)水平的重复测量方差分析(ANOVA)来分析基线校正后波幅总和。条件(P<0.003)和半球(P<0.015)表现出主效应,左半球的反应大于右半球;未发现交互作用(P=0.31)。事后比较(Bonferroni)显示,PF-W和PF-PW没有差异(P>0.99),两种情况下的反应都明显强于PW-W条件(P<0.01),后者没有发现明显新异刺激反应(即所有谐波的Z<1.64)。
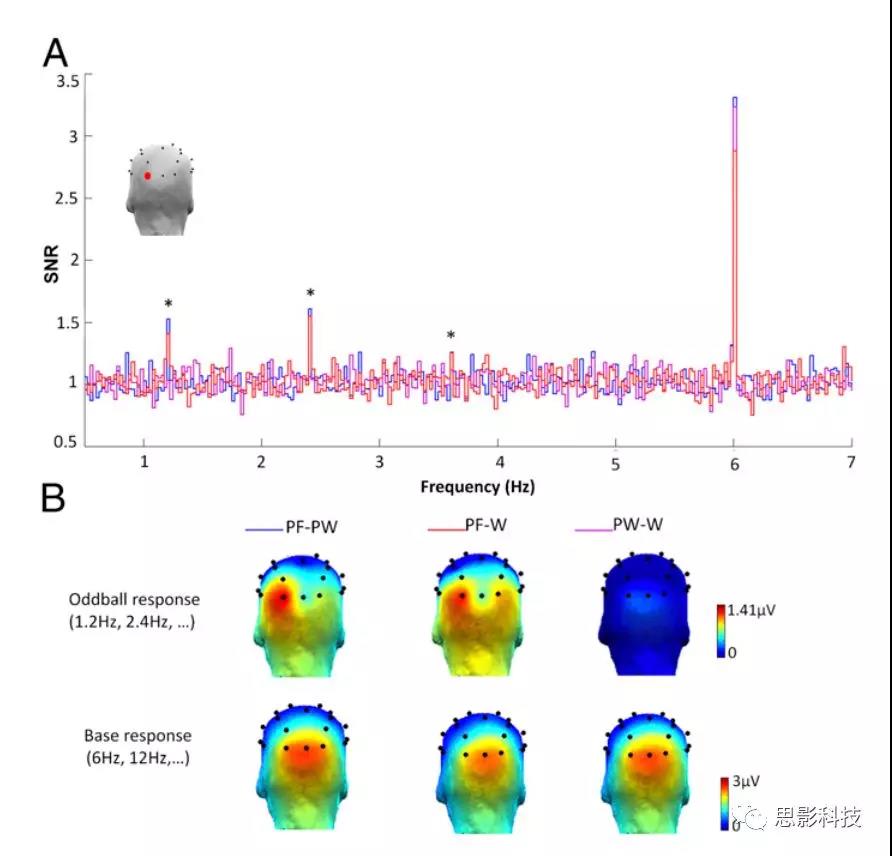
图2 新异刺激频率(1.2Hz,2.4Hz,3.6Hz,4.8Hz)和标准刺激频率(6Hz)的总平均脑电频谱信噪比和头皮地形(n=40)。(A)三种情况下,不同刺激频率下O1(左颞枕电极,地形图上的红点)的信噪比。在1.2hz、2.4hz和3.6hz(*)时,该通道对单词和伪单词的新异刺激反应显著。条件PF-W(红色)和PF-PW(蓝色)没有差异,PW-W(紫色)没有反应。对字符串(PW和W)的反应在左半球(O1)更大。(B)标准刺激频率和新异刺激频率下的每个条件下的地形图。地形图通过显著谐波的波幅减去基线波幅进行校正。
前阅读能力与字符串敏感度的关系
将两个字符串条件合并(“pseudofontf letters”,PF letters),并计算偏侧化指数[(R-L)/(R+L)]。有两种行为测量结果与O1 PF字符新异反应显著相关:字形-音素识别分数(即听到音素时指向正确的字符)(Spearman ρ=0.505;P<0.0001)和字形-音素产出分数(即朗读)(Spearman ρ=0.364;P<0.011)(图3A)。这两种前阅读能力测量指标(GP识别和产出)与O2 EEG 反应(分别为Spearman ρ:0.128,P=0.213;0.016,P=0.462),和与 Oz (Spearman ρ:-0.10;P=0.27,P=-0.157;P=0.16)或 O1(Spearman ρ=-0.093;P=0.285;-0.194;P=0.109)6Hz和其谐波均不相关。综合评分(识别和产出的平均值)与O1反应(Spearman ρ=0.49;P<0.001)以及偏侧化指数(ρ=-0.32;P<0.01)相关,但与O2反应(ρ=0.05;P=0.376)无关。其他语音处理或词汇的测量结果与O1的oddball反应(语音处理:Spearmanρ=0.138;P=0.19;词汇:Spearman ρ=0.101;P=0.259)没有相关性,但字形-音素(GP)的识别和产出分数与语音意识(Spearman ρ=0.463,P<0.001;SpearmanP=0.394;P<0.005)相关。
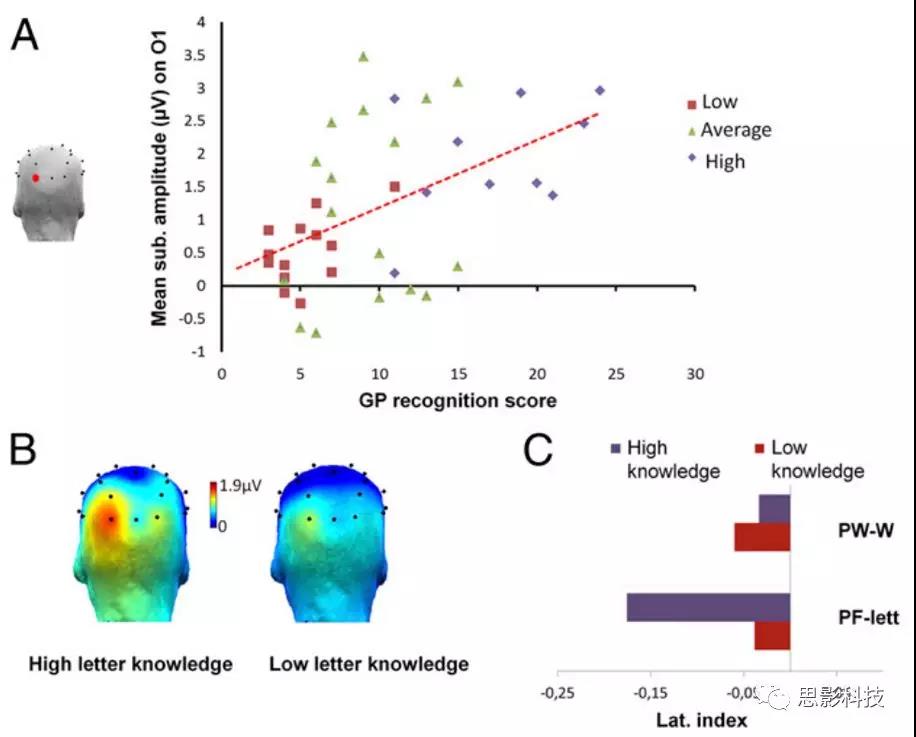
图3。个体表现和脑电反应波幅的关系。(A)用于字符识别的O1平均波幅与字形-音素识别表现呈正相关。(B)具有低水平和较高水平字符认知的儿童对伪字体(PF-letters)中字符的辨别反应(基线校正后)地形图。(C)偏侧化指数(LI),PF-letters条件下,LI在不同组表现不同。在高字符认知组中,LI仅在PF条件下存在显著作用。
视觉熟悉度的影响
在实验2中,用4(实验条件:PF-letters, visual control (VC)-symbols, VC-script,VC-drawings;图4]×2(半球:O1,O2)水平)重复测量方差分析分析基线校正后波幅差异。实验条件(P<0.008)有显著的主效应,半球(F(1,25)<1)无主效应,且这两个因素之间有高度显著的交互作用(P<0.005)。在所有条件下,O1和O2下的反应与0(P<0.0001)显著不同,除了symbol条件下O1[t(25)=1.08;P=0.29]和VC-script条件下两电极(P>0.06)反应(图4和)。在VC-drawings条件下,O1和O2之间没有差异[2.06μV vs. 2.09μV;t(25)=-0.13;P=0.89]。对于PF-letter,左半球的反应强于右半球[LH:1.64μV vs.RH:1.24μV;t(25)=2.07;P<0.04]。对于VC-symbol条件,右半球的反应更强(RH:0.89μV vs.LH:0.27μV;t(25)=-2.80;P=0.010),而VC-script条件下没有明显的偏侧化(t(25)=0.60;P=0.5)。
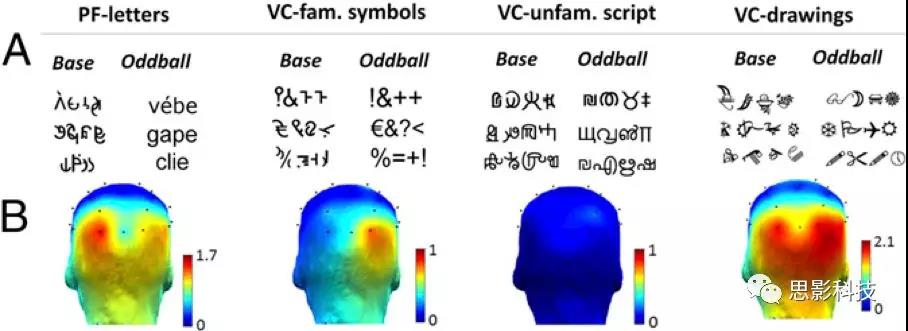
图4。刺激特征和地形图举例(实验2)。(A)标准和新异刺激,以及每个条件的地形图。(B)(1.pseudofont-letters,2. visual control-familiar symbols, 3. visual control-unfamiliar script, and 4.visual control-line drawings)。地形图表示letters, symbols, script, andline drawings的辨别反应,相对于各自的视觉对照刺激(基线校正)。
讨论
通过2分钟的记录,我们在前阅读儿童群体中得到了一个清晰的字符串左半球后侧偏侧化反应。这种反应的幅度因个体而异,并与通过字形-音素识别和产出来测量的前阅读能力相关。这些发现与成人左半球后侧在处理字符串方面的优势一致,成人结果基于多种研究方法包括这里使用的范式。然而,本研究挑战了这样一种观点,即左半球的特异化在自然环境中出现得相对较晚:经过至少1年的正式阅读教学才会出现左半球特异化。在理论层面上,这一发现表明,在发育过程中,大脑后部视觉区和前侧语音区之间建立联系的快速可塑性。从社会的角度来看,它通过具有高度灵敏性、客观性且不需要明确行为反应的研究方法,为阅读习得的研究开辟了新的视角。
用一种灵敏方法证明左半球早期偏侧化
据我们所知,先前的研究并没有发现,在没有正规教育或强化和长期训练的学龄前儿童中,字符串加工的左半球优势。一项功能磁共振成像研究发现,4-5岁儿童对单个符号(字符和数字)的左梭形/颞下叶反应大于单个物体,但此研究没有探索假字符。另一项研究发现左梭状回对单个字符的反应比假字符和形状大,但在右前梭状回也发现了类似的反应,并且与大脑半球的相互作用没有得到验证。而且,因为一个字符可能无法起到类似于字符串的语言处理作用,因此单个字符和字符串可能不会被相同的神经结构处理。在视觉运动和字形-音素训练之后,学龄前儿童对单个字符或甚至字符串的EEG和/或fMRI反应也可能比对符号反应有所增加。然而,在这些研究中,没有对半球偏侧化进行统计学检验或没有达到显著性,倾向性反应可能在几周内消失。此外,到目前为止,字符认知与学龄前儿童大脑半球特异化之间的关系还不清楚。尽管在具有高字符认知的前阅读儿童中发现了右半球N1文本倾向,但也发现了相反的发现(即与阅读结果差相关的右半球N1增强),反应倾向与字符认知之间没有关系。
本研究在学龄前儿童的电生理反应中观察到的高度显著的左半球偏侧化反应可能是由于本方法的几个关键方面。首先,每个字符串都被伪字体前后屏蔽。因此,感兴趣的EEG反应反映了字符串和伪字体之间的非差异性固有对比。所使用的伪字体的数量和广泛多样性,以及它们的大小变化,实际上消除了低级提示对字符串的特定响应的贡献。此外,快速的呈现率(即,每串略低于166毫秒)迫使人们一眼检测字符串,进一步将这一过程与一般的感知和认知功能分离开来。我们注意到,类似的逻辑已经在“快速刺激流范式”中使用,从而产生了一种大约出现在4Hz的“识别电位”(RP),用于表征在各种视觉“背景”刺激中插入的目标刺激。这个RP在成人中左偏,并且随着对比刺激所要求的辨别水平的增加而增加。在目前的频域分析方法中,通过将确定反应频率范围而不是将其扩展到宽带频率ERP成分中显著增强SNR(即信噪比)。
文本材料早期左半球偏侧化:刺激特性或功能加工
阅读前儿童对文本的左偏反应与同样范式的成人反应模式一致。尽管两组之间没有直接的比较,但是儿童的辨别反应似乎出现在比成人的颞枕反应更为偏后的区域。在fMRI中也观察到了类似的发育差异,其中儿童对单词的敏感度在后脑区比成人高。这种效应被认为反映了在整个单词识别变得自动化前,儿童更依赖视觉单词形式系统(VWFS)的后部加工。由于目前的FPVS-EEG方法不需要对单词进行明确的处理,因此受动机和理解以及注意和决策过程的影响较小或不受影响,因此可以用发展过程中完全相同的测量方法相对容易地检验这一假设。
字符辨别的神经测量与前阅读能力行为测量之间的关联支持这样的观点:这些特定的视觉模式与儿童字形-音素关联加工相关。事实上,在婴儿的大脑边缘区域,语音回路已经出现左半球偏侧化。因此,在阅读习得过程中,字符和音素之间的映射,或正字法-音位结合,可以激活左半球同侧大脑后区对字符串的反应。这种观点受到大脑半球内与半球间连接性的支持,和语言区优先和/或预先存在连通性的支持,如功能磁共振成像和弥散张量成像的功能连通性分析。
重要的是,我们第二个实验结果排除了另一种可能性:字符串左半球特异化加工是基于一种普遍的视觉机制:即任何在‘线段连接、中心凹位置和高空间频率’方面具有相似特征的文本的左半球特异化。此外,至少由于三个原因,这些发现还表明,字符串的左半球特异化并不是由于视觉加工仅仅跟踪具有不同低水平特性的两类高空间频率刺激之间的转换。首先,辨别反应只出现在至少知道几个字符的儿童身上。此外,这种左偏反应取决于刺激物的性质,而不是字符状的符号。最后,虽然字符认知能力较好的儿童可能更容易接触到文本,但这种单纯的视觉接触不能解释左半球的特异化。事实上,我们的发现表明,视觉上熟悉的符号与语音表征无关,而是优先激活右半球相关区域,这可能是由于右半球在精通/熟悉视觉形状方面存在优势。
在理论层面上,这一发现至少对我们理解大脑环路发展做出了两个重要贡献。首先,由于最小的字形音素关联知识足以诱发神经变化,这意味着在早期发育过程中建立大脑后部视觉区域和前部音位区域之间的功能连接,可能依赖于先前存在的结构连接性。第二,由于熟悉而非陌生的符号触发(右半球偏侧化的)辨别反应,这表明,对某类视觉刺激的有限认知足以发展用于处理这些刺激的环路。
快速评估(预)阅读能力
除了理论意义之外,我们的研究结果还对发展过程中阅读能力的评估具有潜在的意义。我们能够客观地在80秒的记录中记录字符串敏感度的个体差异,并在第二个儿童样本中复制这个发现。重要的是,FPVS方法不需要任何外显反应,因此允许分离刺激相关和反应相关的过程,这些过程通常在行为测量中相互影响。此外,这种方法能够消除或最小化理解、注意、动机和决策过程等因素的影响,这些因素在个体和发展过程中可能有很大的差异。因此,这种灵敏(即高信噪比)的方法提供了一个强大的工具来快速评估学习阅读的发展。从临床和教育神经科学的角度来看,因为对阅读障碍进行早期干预能够产生更好的结果,在正式教育开始之前发展灵敏的研究方法是非常重要的。诚然,作为一种有效的临床研究工具,该方法仍需进一步改进,进而在所有被试,更多刺激序列和高噪声EEG反应的表现出较高重测一致性(即可靠性)。
综上所述,我们的研究提供了明确证据证明5岁前阅读儿童的左半球后脑区能够进行字符串辨别,并且这种大脑反应的幅度与前阅读能力,即字符认知有显著的联系。这些观察指出,在发育过程中,左半球功能通过早期语音和视觉之间的相互作用形成。
原文:Left corticalspecialization for visual letter strings predicts rudimentary knowledge ofletter-sound association in preschoolers

微信扫码或者长按选择识别关注思影
第十四届磁共振脑网络数据处理班(重庆,7.26-31)
第三十届磁共振脑影像基础班(南京,7.31-8.5)
第十届脑影像机器学习班(南京,6.30-7.5)
第十二届磁共振弥散张量成像数据处理班(南京,6.18-23)
第二十届脑电数据处理中级班(重庆,8.9-14)
如对思影课程感兴趣也可微信号siyingyxf或18983979082咨询。觉得有帮助,给个转发,或许身边的朋友正需要。请直接点击下文文字即可浏览思影科技其他课程及数据处理服务,欢迎报名与咨询,目前全部课程均开放报名,报名后我们会第一时间联系,并保留名额。
小动物磁共振脑影像数据处理班(预报名,南京)
第二十一届脑电数据处理中级班(南京,9.7-12)
第七届眼动数据处理班(南京,7.26-30)
脑电信号数据处理提高班(预报名)
脑磁图(MEG)数据处理学习班(预报名)
思影科技功能磁共振(fMRI)数据处理业务
思影科技弥散加权成像(DWI/dMRI)数据处理
思影科技脑结构磁共振成像数据处理业务(T1)
思影数据处理业务三:ASL数据处理
思影科技脑电机器学习数据处理业务
思影数据处理服务五:近红外脑功能数据处理
思影数据处理服务六:脑磁图(MEG)数据处理
招聘:脑影像数据处理工程师(重庆&南京)
BIOSEMI脑电系统介绍
目镜式功能磁共振刺激系统介绍